subscribe to our mailing list:
|
SECTIONS
|
 |
|
|
Icon of Obfuscation
Jonathan Wells' book Icons of Evolution and why
most of what it teaches about evolution is wrong
Version 1.0
[Last Modified: November 24, 2002]
Posted May 26, 2003
Contents
- Introduction
A
chapter-by-chapter critique of Icons of Evolution:
- Chapter 2:
The Miller-Urey experiment
- Chapter 3:
Darwin's Tree of Life
- Chapter
4: Homology in Vertebrate Limbs
- Chapter 5:
Haeckel's Embryos
- Chapter 6:
Archaeopteryx -- The Missing Link
- Chapter 7: Peppered
Moths
- Chapter 8:
Darwin's Finches
- Chapter 9:
Four-Winged Fruit Flies
- Chapter 10: Fossil
Horses and Directed Evolution
- Chapter 11:
From Ape to Human: The Ultimate Icon
- Conclusions
- Acknowledgements
- References
- Endnotes
Jonathan Wells' book Icons of Evolution:
Science or Myth? Why Much of What We Teach About Evolution Is Wrong
(henceforth Icons) makes a travesty of the notion of honest scholarship.
Purporting to document that "students and the public are being systematically
misinformed about the evidence for evolution," (p. XII)
via common textbook topics such as peppered moths, embryo similarities, and
fossil hominids [2],
Icons in fact contains a bevy of its own errors.
This is not original -- creationists have been making mistakes about evolution
for years. Newly and more insidiously, however, Icons contains numerous
instances of unfair distortions of scientific opinion, generated by the
pseudoscientific tactics of selective citation of scientists and evidence,
quote-mining, and "argumentative sleight-of-hand," the last meaning Wells'
tactic of padding his topical discussions with incessant, biased editorializing.
Wells mixes these ingredients in with a few accurate (but always incomplete)
bits of science and proceeds to string together, often in a logically arbitrary
fashion, a narrative that is carefully crafted to make the semblance of an
honest case for Wells' central defamatory accusation: that mainstream biologists
are "dogmatic Darwinists that misrepresent the truth to keep themselves in
power" (pp. 242-243).
This essay will show that it is Wells' book Icons that is shot through
with misrepresentations.
The central pillar of Wells' case is this:
"Some biologists are aware of difficulties with a particular icon
because it distorts the evidence in their own field. When they read the
scientific literature in their specialty, they can see that the icon is
misleading or downright false. But they may feel that this is just an isolated
problem, especially when they are assured that Darwin's theory is supported by
overwhelming evidence from other fields. If they believe in the fundamental
correctness of Darwinian evolution, they may set aside their misgivings about
the particular icon they know something about." (Icons, pp.
7-8) In other words, Wells argues that the specialists know about
the problems in their field of expertise, but that everyone thinks that the
evidence supporting evolution is somewhere else. This is just plain false, as we
shall see -- the experts in each field have explicitly stated that the
evidence in their field supports evolutionary theory, and further they
have supported their statements with evidentiary arguments. If Wells' contention
about the experts is false, then Wells' argument collapses. Wells likes asking
questions; it is now time for him to answer some.
A key question in origin-of-life research is the
oxidation state of the prebiotic atmosphere (the current best guess is that the
origin of life occurred somewhere around 4.0-3.7 bya (billion years ago)). Wells
wants you to think that there is good evidence for significant amounts free
oxygen in the prebiotic atmosphere (significant amounts of free oxygen make the
atmosphere oxidizing and make Miller-Urey-type experiments fail). He spends
several pages (14-19) on a pseudo-discussion of the oxygen issue, citing sources
from the 1970's and writing that (p. 17) "the controversy has never been
resolved", that "Evidence from early rocks has been inconclusive," and
concluding that the current geological consensus -- that oxygen was merely a
trace gas before approximately 2.5 bya and only began rising after this point --
was due to "Dogma [taking] the place of empirical evidence" (p. 18). None of
this is true (see e.g. Copley, 2001).
-
Certain minerals, such as uraninite, cannot form under significant exposure
to oxygen. Thick deposits of these rocks are found in rocks older than 2.5 bya
years ago, indicating that essentially no oxygen (only trace amounts) was
present. On page 17 Wells notes that uraninite deposits have been found in
more recent rocks, but neglects to mention to his readers that these only
occur under rapid-burial conditions, whereas ancient deposits of uraninite
occur in slow deposition conditions, for example in sediments laid down by
rivers, so that the minerals were exposed to atmospheric gases for significant
periods of time before burial.
-
'Red beds' are geologic features containing highly oxidized iron (rust)
indicative of high amounts of oxygen. Wells (p. 17) notes that red beds are
found before 2 bya, but fails to mention that the temporal limit of red beds
is just a few hundred million years before 2 bya.
-
Wells doesn't even mention the evidence that banded iron formations
(incompletely oxidized iron indicative of ultralow-oxygen conditions) are very
common prior to 2.3 bya and very rare afterwards.
-
Wells also doesn't mention that early paleosols (fossil soils) from about
~2.5 bya contain unoxidized cerium, impossible in an oxygenic atmosphere
(e.g., Murakami et
al., 2001).
-
Finally, Wells doesn't mention to his readers that pyrite, a mineral even
more vulnerable to oxidation than uraninite, is found unoxidized in pre-2.5
bya rocks, and with significant evidence of long surface exposure (i.e. grains
weathered by water erosion; e.g. Rasmussen
and Buick, 1999).
Why does Wells leave out the converging independent lines of geological
evidence pointing to an anoxic early (pre ~2.5 bya) atmosphere?
The famous Miller-Urey experiments used a strongly reducing
atmosphere to produce amino acids. It is important to realize that the original
experiment is famous not so much for the exact mixture used, but for the
unexpected discovery that such a simple experiment could indeed produce crucial
biological compounds; this discovery instigated a huge amount of related
research that continues today.
Now, current geochemical opinion is that the prebiotic atmosphere was not so
strongly reducing as the original Miller-Urey atmosphere, but opinion varies
widely from moderately reducing to neutral. Completely neutral atmospheres would
be bad for Miller-Urey-type experiments, but even a weakly reducing atmosphere
will produce lower but significant amounts of amino acids. In the approximately
two brief pages of text where Wells actually discusses the reducing atmosphere
question (p. 20-22), Wells cites some more 1970's sources and then asserts that
the irrelevance of the Miller-Urey experiment has become a "near-consensus among
geochemists" (p. 21).
-
This statement is misleading. What geochemists agree on is that if
the early earth's mantle was of the same composition as the modern mantle
and if only terrestrial volcanic sources are considered as contributing
to the atmosphere, and if the temperature profile of the early
atmosphere was the same as modern earth (this is relevant to rates of hydrogen
escape) then there will be much less hydrogen compared to Miller's
first atmosphere (20% total atm.). Even if this worst-case scenario is
accepted, hydrogen will not be completely absent, in fact there is a long list
of geochemists that consider hydrogen to have been present (although in lower
amounts, roughly 0.1-1% of the total atmosphere). At these levels of
H2 there is still significant (although much lower) amino acid
production.
-
Also, many geochemists think that these conditions do not represent
the early earth, contrary to the impression given by Wells. For example, on p.
20, Wells mentions terrestrial volcanos emitting neutral gases
(H2O, CO2, N2, and only trace H2),
but he fails to mention that mid-ocean ridge vents could have been
significant sources of reduced gases -- they are important sources of reduced
atmospheric gases even today, emitting about 1% methane (Kasting and
Brown, 1998) and producing reduced hydrogen and hydrogen sulfide (e.g. Kelley et
al., 2001; Perkins, 2001;
Von Damm,
2001) and potentially ammonia prebiotically (Brandes et
al., 1998; Chyba, 1998). Why
does Wells exclude oceanic vents from consideration?
-
Another strange omission is that Wells completely fails to mention the
extraterrestrial evidence, which is the only direct evidence we have of the
kinds of chemical reactions that might have occurred in the early solar
system. For example he neglects to mention the famous Murchison meteorite,
which contains mixtures of organic compounds much like those produced in
Miller-Urey style experiments, and which constitutes direct evidence that just
the right kind of prebiotic chemistry was occurring at least somewhere in the
early solar system, and that some of those products found their way to earth
(see e.g. Engel and
Macko, 2001 for a recent review).
-
Wells asserts that since the 1970's, non-reducing atmospheres have become
the "near-consensus." The latest article that Wells cites supporting this
view, however, is a 1995 nontechnical news article in Science (Cohen, 1995). Why
doesn't he quote Kral et al.
(1998), who write,
The standard theory for the origin of life postulates that life
arose from an abiotically produced soup of organic material (e.g., Miller,
1953; Miller, 1992). The first organism would have therefore been a
heterotroph deriving energy from this existing pool of nutrients. This
theory for the origin of life is not without competitors (for a review of
theories for the origins of life see Davis and McKay, 1996), but has
received considerable support from laboratory experiments in which it has
been demonstrated that biologically relevant organic materials can be easily
synthesized from mildly reducing mixtures of gases (e.g., Chang et
al., 1983). The discovery of organics in comets (e.g., Kissel and
Kruger, 1987), on Titan (e.g., Sagan et al., 1984), elsewhere in the
outer solar system (e.g., Encrenaz, 1986), as well as in the interstellar
medium (e.g., Irvine and Knacke, 1989) has further strengthened the notion
that organic material was abundant prior to the origin of
life.
None of this is meant to convey the impression that no controversies exist
(both Cohen
(1995) and the Davis and McKay
(1996) article cited by the above-quoted Kral et al.
(1998) are about the various competing hypotheses about the origin of life).
But textbooks generally mention some of these hypotheses (briefly of course, as
there is only space for a page or two on this topic in an introductory
textbook), and furthermore generally mention that the original atmosphere was
likely more weakly reducing than the original Miller-Urey experiment
hypothesized, but that many variations with mildly reducing conditions still
produce satisfactory results. This is exactly what is written in the most
popular college biology textbook, Campbell et
al.'s (1999) Biology, for instance. In other words, the textbooks
basically summarize what the recent literature is saying. The original
Miller-Urey experiment, despite its limitations, is also repeatedly cited in
modern scientific literature as a landmark experiment. So why does Wells have a
problem with the textbooks following the literature? Wells wants textbooks to
follow the experts, and it appears that they are.
Wells writes (p. 22) as if the RNA world is an
alternative to failed Miller-Urey-style experimentation. He cites no source for
this claim, because the claim is pure obfuscation.
-
The RNA world hypothesis is complementary, not opposed, to Miller-style
prebiotic syntheses, as it is meant to explain how genetic replication got
going without DNA, several steps down the road after prebiotic syntheses.
-
Wells gives the impression that there are only two possible starts to life
on earth, Urey-Miller style syntheses and the RNA world. Wells misleadingly
cites several quotes that taken alone suggest that the RNA world is
impossible, and that there is no remaining scientific explanations for life on
earth. However, most authorities agree that the RNA world was one stage
of the origin of life, rather than the very first stage, and that it was
proceeded by a pre-RNA world. Indeed, the very authors he quotes to suggest
that the RNA world is impossible go on to explain the concept of a pre-RNA
world and how an RNA world would arise from that, but Wells omits all mention
of this. Wells doesn't bother to cite recent work on precursors to the RNA
world, see for example Cavalier-Smith
(2001) for an introduction and references to ideas on this such as the 'NA
world' and 'lipid world' (for the latter, see e.g. Segre et
al., 2001).
3. Chapter 3: Darwin's Tree of Life
Wells mixes up several issues in this chapter. As we saw in the previous
chapter, he will give several topics each a cursory and incomplete treatment,
raising doubts about each subject and connecting them together whether they are
logically connected or not.
Here Wells is running down a path well-worn by
his creationist and 'designist' (IDist) colleagues. As a result there is already
significant literature available on the "animal phyla appeared suddenly, and
without precursors, and all equally far apart from each other"-sort of
contention.
-
See for example Conway Morris (Conway Morris,
1998) for an authoritative discussion, and Knoll and
Carroll (1999) which is freely available online. A particularly
interesting perspective is that of Keith B. Miller, geologist, evolutionist
and evangelical Christian, who wrote an article (1999) in Perspectives on
Science and Christian Faith entitled ( "The Precambrian to Cambrian Fossil
Record and Transitional Forms," in which he
wrote,
There is much confusion in the popularized literature about the
evidence for macroevolutionary change in the fossil record. Unfortunately,
the discussion of evolution within the Christian community has been greatly
influenced by inaccurate presentations of the fossil data and of the methods
of classification. Widely read critiques of evolution, such as Evolution:
A Theory in Crisis by Denton, and Darwin on Trial by Johnson,
contain serious misrepresentations of the available fossil evidence for
macroevolutionary transitions and of the science of evolutionary
paleontology. [...] The implication of much of the evangelical Christian
commentary on macroevolution is that the major taxonomic groups of living
things remain clearly distinct entities throughout their history, and were
as morphologically distinct from each other at their first appearance as
they are today. There is a clear interest in showing the history of life as
discontinuous, and any suggestion of transition in the fossil record is met
with great skepticism. The purpose of this short communication is to dispel
some of these misconceptions about the nature and interpretation of the
fossil record.
-
On page 38 of Icons Wells quotes page 30 of Simon Conway Morris'
(1998) book The Crucible of Creation about the "sharp demarcation"
at the Cambrian explosion. But Conway Morris writes on the very next page
(32-3), "The term 'explosion' should not be taken too literally, but in terms
of evolution it is still very dramatic. What it means is the rapid
diversification of animal life. 'Rapid' in this case means a few millions of
years, rather than the tens or even hundreds of millions of years that are
more typical when we consider evolution in the fossil record." And of course
one of the central points of Conway Morris' entire book is that the
morphological disparity that emerges in the Cambrian, often invoked by Wells
& co. via quotation of people like Stephen J. Gould, is not quite as
radical as the IDists or even Gould would have it. There is a significant
problem in asserting that the animal phyla of the Cambrian are as
morphologically diverse as the fauna today -- in large part this perception is
due to the somewhat arbitrary concepts of 'phylum' and 'body plan'. Conway
Morris writes (p. 170) that "the strangeness of the problematic Cambrian
animals is really a human artifact, a construct of our imagination." On pages
185-195 he uses the fossils Wiwaxia and Halkieria as
transitionals between phyla (which by Wells' account shouldn't
exist) to connect three very 'disparate' major phyla -- Mollusca, Brachiopoda,
and Annelida -- to each other.
-
For someone who self-righteously passes judgment on figures in textbooks,
Wells' figures are atrocious. His Figure 3.4, "Actual Fossil Records of the
Major Living Animal Phyla" purports to show when the various animal phyla
appear in the fossil record. Note that, again, Wells cites no source in his
notes to figure out where his figures came from. However, examining a similar graphic
at the online University of California Museum of Paleontology (UCMP, 2000), we see
that Wells has left a number of phyla out, namely the dozen or so that have
either no fossil record or appear very late relative to the Cambrian phyla.
Notably all of the missing phyla are soft-bodied, but if Wells admitted that
numerous phyla existed without any fossil record whatsoever, he would severely
weaken his argument on page 44 that any small, soft-bodied ancestors of the
animal phyla would have been fossilized if they existed. Wells' figure also
places Rotifera and Phoronida as having fossil records in the Cambrian, which
could be correct if the UCMP page is out of date, but as Wells gives no
references for the figure it is impossible to check.
Wells' second argument against the Tree of Life
deals with the 'molecular clock' hypothesis -- namely that DNA or protein
sequence divergence is regular enough to date ancient splits between lineages.
This hypothesis is indeed being questioned by scientists, as the influence of
things like natural selection may well alter the rate of sequence change (e.g.
cone snail venoms are a fantastic example of rapid sequence divergence under
selective pressure; see Espiritu et
al., 2001). And if these changes occur often enough then getting
accurate clock dates, particularly for distant events, will be very hard. This
is an entirely different thing from determining molecular phylogenies, however,
which is what Wells is actually trying to debunk. But unfortunately for Wells,
there is considerable evidence that these phylogenies are reliable and in
reasonably good accord with phylogenies generated from other data. On the
general subject of accuracy in molecular phylogenies see Theobald 2002b
and for recent work on phyla evolution and metazoan molecular phylogenies, which
are quite certainly not in crisis, see recent articles in places Evolution
and Development (e.g., Collins
and Valentine, 2001; Peterson
and Eernisse, 2001).
-
Wells' Figure 3.6 (p. 47) is laughable. He represents molecular phylogenies
as comparing four (count 'em) base pairs among three organisms.
It looks like this:
|
DNA Sequence |
| Organism 1 |
ATCG |
| Organism 2 |
ATCT |
| Organism 3 |
ATGT |
Why couldn't Wells at least get a real dataset, like any
article or even textbook would use? You have to compare at least a few dozen
base pairs before you can see the uncanny way that organisms in the same genus
match up far better than organisms in different classes (for example). Here,
for example, is an alignment of some
cytochrome C amino acid sequences from various organisms (for discussion
see here). If Wells were interested in giving his readers a useful
graphic, he could have easily found something like this, published in a 1992
article of the Journal of Molecular Evolution:
Ape-human relationships. The following example comes from the
mitochondrial DNA sequence data from Horai et al. (1992, Journal of Molecular
Evolution 35: 32-43). Just small blocks of data are shown from three of the
several protein-coding and tRNA loci
analyzed (COI =
cytochrome oxidase I locus).
From locus: COI tRNALys. ATPase 8
position 2664 2671 4026 4036 4410 4418
| | | | | |
Human ACACCATA ACTTTCACCGC AAAAAATTA
Chimp ACACCATA ACTTTCACCGC AAAAAACTA
Bonobo ACACCATA ACTTTCACCGC AAAAAACTA
Gorilla CCACCACA ACATTCACCGC AAAAAACTT
Orangutan CCACCACA ACATTCACTGC AAAACCCCA
Gibbon CCACCATA ACATTCACCGC TAAACCCCA
|
(Source for sequences and caption: http://www.homepage.montana.edu/~mlavin/b403/lec12.htm.
See that page, part of notes for a course on evolution at Montana State, for
further discussion.) A discussion of the sequence analysis
and the mathematics of nested phylogenies is here: www.talkorigins.org/faqs/comdesc/section1.html)
-
Wells engages in a bit more quote-mining on page 51, picking some molecular
phylogenies studies that seem ridiculous.
Even when different molecules can be combined to give a single
tree, the result is often bizarre: A 1996 study using 88 protein sequences
grouped rabbits with primates instead of rodents; a 1998 analysis of 13
genes in 19 animal species placed sea urchins among the chordates; and
another 1998 study based on 12 proteins put cows closer to whales than to
horses.
What Wells isn't telling you is that some of these results are not in fact
ridiculous.
-
Cows, for example, are artiodactyls which are indeed thought to be
closely related to whales, a suspicion which has received striking
confirmation from recent transitional fossil discoveries (see the webpage of
the discoverer Thewissen).
-
Sea urchins (phylum echinodermata) do indeed group "among the chordates"
but this is because they are a sister group to chordates, not within
chordates as Wells implies. This taxonomy is a long accepted fact (see e.g.
the Metazoa
page of the Tree of Life, especially "D" for "deuterostomes," the group
which includes echinoderms and chordates but excludes moulting organisms
like arthropods). The very paper that Wells cites recognizes these
distinctions explicitly.
-
The rabbits and rodents study, on the other hand, has methodological
flaws (although the two groups are indeed more distantly related than the
nonexpert might expect). All of this is discussed in detail, with
references, by talkorigins
poster John Harshman.
The 'Tree of Life' is the idea, most
famously advocated by Darwin, that all known life is descended from a common
ancestor and is connected by a phylogenetic 'tree':
"The affinities of all the beings of the same class have sometimes been
represented by a great tree. I believe this simile largely speaks the truth.
The green and budding twigs may represent existing species; and those produced
during each former year may represent the long succession of extinct species.
... As buds give rise by growth to fresh buds, and these, if vigorous, branch
out and overtop on all sides many a feebler branch, so by generation I believe
it has been with the great Tree of Life, which fills with its dead and broken
branches the crust of the earth, and covers the surface with its ever
branching and beautiful ramifications." (Darwin, Origin of
Species)
This idea has recently been implemented on the web in a splendid fashion. See
the Tree of Life Web Project
Home Page.
In Icons, Wells has a ball with recent scientific debates over whether
or not lateral gene transfer mixed up ancient genomes so much that deepest
branches of the tree are mixed up. Basically, some scientists have proposed that
the idea of a single "last common ancestor" should be replaced with the idea of
a "last common gene pool" that the extant three domains of of life --
eukaryotes, archaea and eubacteria, in one classification scheme -- gradually
emerged from. Carl
Zimmer (2001) describes this as the 'Mangrove of Life' idea. Wells (of
course) milks this for all it's worth, proclaiming the downfall of common
descent and the 'uprooting of the tree' and whatnot, but he is distorting
things.
- This entire debate is, among scientists, about the very oldest part of the
tree, known as the 'root.'
This is where the lineages of the three fundamental 'domains' of life come together.
Apart from being the most remote event to study timewise, the question of the
rooting of the tree is greatly complicated by lateral gene transfer, by
differing rates of evolution between genes and lineages, by the fact that
eukaryotes are the result of symbioses between archaea and eubacteria, and by
the fact that, by definition, the Tree of Life has no outgroup, which creates
technical problems for placing the root. Scientists are attempting to discern
the most ancient events in the history of life here, so complications are to
be expected. One recent article that is highly skeptical of much of the work
that Wells cites is Cavalier-Smith
(2002).
-
What Wells does not point out is that this entire controversy has precious
little to do with eukaryote phylogeny (which is coming along just fine, thank
you, see e.g. Baldauf et
al., 2000), and nothing to do with metazoan phylogeny (previously
discussed), all of which will remain perfectly traditional and tree-like no
matter who wins the debate over rooting the tree. To see what this means, go
to the Tree of Life webpage (http://tolweb.org/), if you click on the tree, you will find that the
first page lists the three domains. This is what the tree vs. mangrove debate
is about. If you click on eukaryotes,
you have entered the zone where, regardless of the debate outcome, it appears
that phylogenies will remain tree-like -- in other words, essentially the
entire mapped tree. Wells' Figure 3.8 (p. 53) is very misleading in this
respect -- all macroscopic life, and a great deal of microscopic life, fits
into the tree branching off from the 'molecular thicket'.
-
How any of this is supposed to call evolution into question is not at all
clear -- Wells certainly has not proposed a model that explains things better.
And the fact that textbooks aren't completely up to speed on current
scientific debates is not only not surprising, it's the way things should be.
This particular controversy is far from resolved, and until it is, there is no
real advantage of putting it into textbooks. Even if the 'mangrove' model is
eventually accepted, it is rather difficult to see how this makes "descent
with modification from common ancestors...not even a well-supported theory"
(Icons, p. 58), as the debate will have been resolved by figuring out
what the true lines of descent are. If it turns out our remotest ancestors are
a community of gene-trading bacteria rather than a single one (and it should
be remembered that it is also possible that a community of gene-trading
bacteria could still be descended from one bacteria), then this will be
significant but hardly something that overthrows the evolutionary view of
life. And as mentioned before, there are good reasons to think that the
traditional tree model will basically work even down near the root. E.g.,
Cavalier-Smith (2002) writes,
"Recent worries that lateral transfer is so rampant (Doolittle, 1999a, b)
that we may never reconstruct organismal trees are certainly false for
eukaryotes and probably incorrect for bacteria. I agree with Doolittle
(2000) that the widely accepted tree needs uprooting, not because of lateral
transfer, which is not seriously confusing with respect to the root, but
because quantum evolution caused misrooting of the paralogue tree. We may
safely replant it as shown in Figs 1, 2 and 7. The idea that genome
composition in the cenancestor was so fluid (Woese, 1998, 2000) that we
cannot use cladistic arguments to reconstruct it is more profoundly
mistaken, being based on the basic misinterpretations of the universal tree
and the evolutionary significance and timing of the differences between
eubacteria and neomura explained above. It has long been clear
(Cavalier-Smith, 1981, 1987a, b, 2001) that the cenancestor was a normal
eubacterium, not a progenote. As Woese (1998, 2000) and Doolittle recognize,
the most readily transferred genes are not a random sample of the whole, but
obey certain rules (Rivera et al., 1998; Jain et al., 1999;
Martin, 1999)." (Cavalier-Smith,
2002, p. 62)
On the last page of this chapter of
Icons (p. 58), Wells recounts his story of an unnamed Chinese
paleontologist who visited the U.S. in 1999 and who Wells quotes as saying, "In
China we can criticize Darwin, but not the government; in America, you can
criticize the government, but not Darwin." Given past experience with
antievolutionist anecdotes, it is to be strongly suspected that there is more to
this story than Wells is telling us.
-
This is particularly true if one reads the account of Nigel Hughes
(Department of Earth Sciences, University of California, Riverside) of a small
symposium held in June 1999 in China, where many well-preserved early metazoan
fossils have recently been discovered. Hughes says that the meeting, the
"International Symposium on the Origin of Animal Body Plans and Their Fossil
Records," was organized by Professor Chen Jun-yuan -- although, as we will see
below, it appears that the Discovery Institute played a significant role as
well. Hughes was one of about 50 symposium participants.
Hughes reports on the meeting in the March/April 2000 issue of the journal
Evolution and
Development (Vol. 2, Issue 2, pp. 63-66). Most of the report is
typical scientific meeting details, but Hughes takes the last several
paragraphs of his report to discuss "the most curious aspect of the
meeting":
The most curious aspect of the meeting, and the most embarrassing for
Western scientists (particularly those from the United States), was the
presence of individuals supported by the Discovery Institute -- a
Seattle-based foundation that proclaims intelligent design as a scientific
explanation for biological diversity. The involvement of the Institute came
as a surprise to the more conventional attendees, especially when it became
obvious that the Institute had played a key role in the organization of the
conference, unbeknownst to the scientific community. Several talks were
presented along this theme, the main thesis of which seemed to be the old
Pallian arguments wrapped in a variety of molecular guises. Michael Denton
spoke on what he saw as a failure of genetics to unveil a universal
explanation for biological form, Paul Nelson on maternal effect genes, and
Jonathan Wells on homeotic genes. It takes guts to expose yourself in this
manner to a generally incredulous audience, but it also places special
demands if science is your objective. I was depressed to find that my
rudimentary understanding of molecular biology was sufficient to spot
egregious errors, candidly dispatched by Eric Davidson. Wells's claim that
aspects of Hox gene control, instead of providing yet more evidence
for homology and common ancestry, actually suggest that all metazoan phyla
arose independently gives the flavor of what was offered. In doing so he
effectively denied any defensible meaning in the words such as deuterosome
or ecdysozoan, well established higher taxa which have been erected on
characters other than those genes that influence segment identity. A bold
claim, but one he could not reasonably defend as questioning revealed.
Denton was dismayed that biotic systems are more complicated than some
geneticists had expected in the 1960s, but the logical connection between
this and his belief in immutable natural designs was left unexplained. And
so it went on. The only thing new here was the presence of these arguments
at a meeting that was ostensibly billed as being scientific.
How does one deal with such situations? Those speaking were accompanied
by a coterie of supporters, including a "cosmic reporter" and one-by-one
scientists attending courteously answered their questions. Many of us,
myself included, reluctantly agree to be interviewed on tape. As guests in
China a major public blow-up was to be avoided, but looking back I wish I
had been more aggressive. We are all used to arguing science, but we are not
used to telling people that we suspect their motives. Perhaps we have to
become so, because the extent, if any, to which Chinese colleagues had been
made aware of the controversial nature of the Discovery Institute, and its
political agenda within the United States, remained unclear. What was clear
is that the Discovery Institute is actively encouraging Chinese scientists,
by means of funding, to promote a view of the Chengjiang fauna to which they
are sympathetic.
Several Chinese scientists gave presentations that emphasized the sudden
appearance of phyla, hinting at the need for a new "top-down" mechanism of
evolution -- music, of course, to creationist ears. Although the Chengjiang
fauna does forcefully remind us that many body plans were firmly established
by early in the Cambrian, it does little more than focus attention on the
interesting things that happened around the Precambrian/Cambrian boundary.
The "phylogenetic lawn" idea is hardly new (recall, for example, Gould's
Wonderful Life), and is clearly an inaccurate view. Given the
generous way in which scientists at the meeting explained this and other
matters to those allied with the Discovery Institute it is disappointing to
find commentaries in the Wall Street Journal (August 16, 1999)
proclaiming that Chinese scientists have new evidence that questions the
very basis of evolution. Predictably enough, the Discovery Institute turns
out to be uninterested in scientific rigor, and they will do whatever it
takes to promote their agenda, including taking advantage of Chinese
scholars. Creationism is not only a specter that haunts rationality in the
United States, but it is also willing to employ a little cultural
imperialism if it furthers the cause.
(Hughes, 2000, "The rocky road to Mendel's play," Evolution and
Development, 2(2), 63-66)
-
Wells discusses his story further in the footnotes (Icons, p. 278),
and implies that revealing the name of the paleontologist would expose the
scientist to persecution from dogmatic Darwinists. But this is nonsense, as it
seems certain that the Chinese paleontologist was at the above symposium
(quite possibly he was the organizer, although I have no further evidence for
this than the Hughes piece above), and was clearly given a chance to give his
views. Antievolutionists of course often will characterize mere disagreement
as persecution, but that is their problem.
-
Wells cites the supposed hounding of Colin Patterson as support for Wells'
persecution paranoia. This is a famous creationist canard that has been dealt
with many times before (see e.g. the Colin Patterson FAQ).
Fortunately Robert Hagen (University of Kansas, Department of Ecology and
Evolutionary Biology) has already dealt with Wells' specific claims on
Patterson and given permission to have his analysis quoted here:
The "Darwinist Conspiracy": Wells vs. Reality
The strangest aspect of the book is the bizarre view of mainstream
science Wells presents: The idea that a secret gang of "Darwinists" controls
the teaching of evolution and uses coercion and deceit to suppress all
disagreement! Does Wells present any evidence to support his claim of a
Darwinist conspiracy that mercilessly persecutes any scientist who dares
criticize the dogma?
No. But he does have an anecdote.
On p. 58, he quotes an unnamed Chinese paleontologist as saying: "In
China we can criticize Darwin, but not the government; in America, you can
criticize the government, but not Darwin." Wells expands on this story in
the Research Notes:
"The Chinese paleontologist story has been making the rounds
since I first told it to some colleagues in 1999. Sadly, the principal
reaction from dogmatic American Darwinists has been to demand his name. I
refuse to give it to them, knowing what their colleagues have been doing
to critics since at least 1981, when British paleontologist Colin
Patterson, in a famous lecture at the American Museum of Natural History
in New York, openly questioned whether there is any evidence for
evolution. Afterwards, dogmatic Darwinists hounded him relentlessly, and
Patterson never again voiced his skepticism in public. I fear they would
do the same to the Chinese paleontologist in my story, an excellent
scientist who deserves to be protected from heresy hunters."
(Icons, p. 278)
This would be a compelling story, if it were true. However, in Colin
Patterson's own words, it was hardly "dogmatic Darwinists" who hounded
him:
"Because creationists lack scientific research or evidence to
support such theories as a young earth (10,000 years old), a world-wide
flood (Noah's), and separate ancestry for humans and apes, their common
tactic is to attack evolution by hunting out debate or dissent among
evolutionary biologists. When I published the first edition of this book I
was hardly aware of creationism but, during the 1980s, like many other
biologists I learned that one should think carefully about candour in
argument (in publications, lectures, or correspondence) in case one was
furnishing creationist campaigners with ammunition in the form of
'quotable quotes', often taken out of context." (C. Patterson,
Evolution, 2nd edition, p. 122)
Patterson, who died in 1998, was not an individual who could be cowed
into silence by "dogmatists" of any sort. With regard to the aspect of
evolutionary biology that Wells opposes -- common descent -- Patterson goes
on to state his view clearly:
"I see the general historical theory, common descent, as being
as firmly established as just about anything else in history. We have
compelling reasons to believe that Napoleon and the Roman empire existed,
although we don't know every detail of what went on in Napoleon's life or
in Rome and its colonies; it is much the same with evolution. There is
abundant documentary evidence for Napoleon and the Roman empire; there is
abundant evidence for common descent in the hierarchy of homologies at
both the structural and morphological level, though those documents may
not be so easy to read." (Patterson, p. 123)
The real reason that "experts on the evidence," such as Patterson, don't
reject Darwin's theory is simply that no one -- including Wells -- has found
a better theory:
"...today's theory of evolution is unlikely to be the whole
truth. It is essential to keep in mind the distinction between the general
theory -- evolution has occurred and species are related by descent -- and
theories of mechanism -- natural selection, neutralism, etc. Today's
theory, accepting that evolution has occurred and explaining it by
neo-Darwinism plus neutralism, is the best that we have. It is a fruitful
theory, a stimulus to thought and research, and we should accept it until
nature prompts someone to think of one that is better or more complete."
(Patterson, p. 120)
(Robert Hagen, "Icons of Trouble," The Link: Newsletter of Kansas
Citizens for Science, 1(3), December,
2001)
Is the definition of homology circular?
Wells spends this entire
chapter thoroughly confused about homology, and does his best to confuse his
readers as well. About five minutes of research by yours truly turned up a
perfectly reasonable discussion of homology (Amundson, 2001) which nicely
straightens things out: in a nutshell, homology is detailed similarity of
organization that is functionally unnecessary, meaning the
similarity is unnecessary (the trait in question may be, and usually is,
functional).
This is not a drastically complicated idea, but Wells is able to confuse
himself because post-Darwin, homology is usually defined as 'A feature in two or
more taxa is homologous when it is derived from the same (or a corresponding)
feature of their common ancestor.' Wells accuses scientists of making the
definition circular and assuming common descent rather than providing evidence
for it, but of course if you doubt common descent then you have to go back to
the original definition, wherein you have peculiar similarities that are
functionally unnecessary staring you in the face, begging you for an
explanation. Amundson (2001)
critiques the circularity charge well:
This criticism relies on a faulty view of what scientific
definition amounts to. A scientific definition is not a semantic stipulation
that creates an analytically true statement (i.e. a statement the denial of
which is self-contradictory). Rather, a scientific definition typically states
a property that is considered to be the most deeply explanatory of the
phenomena that are central to the term being defined. Lankester and Mayr
consider ancestry to explain patterns of homology, and stress that fact by
making it the definition. The historical definition is not viciously circular
as long as homologies can be recognized and picked out by criteria other than
common ancestry. It is an empirical fact that homologies (as picked out by the
criteria below) are arranged among organisms in a pattern that is explainable
by common ancestry, and independent evidence from various fields supports
common ancestry as a historical fact.
On pp. 72-78,
Icons cites Rudolf Raff and other scientists in an attempt to build a
case that the use of homology is somehow in crisis. The reality of most of these
cases is that scientists are simply discovering that different kinds of homology
are found at different phylogenetic and organizational levels. Over the last few
years, the growing synthesis of evolutionary biology and developmental biology
has created a hot new subfield referred to as "evo-devo." For an introduction to
evo-devo, see the article "The evolution of evo-devo
biology," which heads up an entire special issue on
the topic that was published freely on the web by the journal Proceedings of
the National Academy of Sciences (PNAS).
The evo-devo specialist Rudolph Raff recently (Nov.-Dec. 2001) wrote an
editorial in the journal Evolution and Development entitled "The
creationist abuse of evo-devo," specifically on Wells and his book Icons.
Raff writes, in part,
Icons of Evolution presents the dark view of
evolutionary biologists held by Wells. He says that we are involved in a
conspiracy to consciously lie in what we teach students and present in our
writings. Claims of deliberate scientific fraud and "Darwinian censorship"
reaches a crescendo as the book progresses. These are strong accusations built
on a shaky scaffolding of special pleading and deceptive use of quotations.
[...] Wells notes correctly that there is not a necessary connection between
homologous genes and homologous structures, nor must homologous structures
arise from similar developmental processes. [Wells and colleagues conclude
that...] "naturalistic mechanisms proposed to explain homology do not fit the
evidence." What logical gymnastics! If it is unexplained, it must be
unexplainable by evolutionary biology. If it's be unexplainable by
evolutionary biology, it must require an intelligent designer. Unfortunately,
as the influence of the intelligent designer grows in this train of thought,
the relationships between phenomena and explanations becomes increasingly
arbitrary. Finally one reaches a point where all biological features are
"special creations" and other explanations become unnecessary. (Raff,
2001)
For a very detailed introduction to dozens of detailed homologies (none
mentioned by Wells except the vague 'similarity' idea) within the chordates that
have been discovered via comparative biology, see this webpage on Chordate
Anatomy and Evolution).
5. Chapter 5: Haeckel's Embryos
In the interests of forthrightness, one point must be conceded straight out:
Haeckel's embryo drawings have no place in textbooks except as an example of how
erroneous ideas can get tacked onto important truths and perpetuated even after
being debunked (Haeckel's inaccurate drawings have actually been 'exposed'
multiple times since the 1800's, the Richardson
et al. (1997) article that Wells cites being only the most recent
example). However, Wells as usual exaggerates the implications of this for
evolution.
- . In his critique of Wells,
the aforementioned Raff wrote,
Richardson et al. (1997) showed that Haeckel falsified
the degree of external appearance of these embryos to exaggerate similarity
of phylotypic stage. To Wells this means that "scientists have long known
that drawings showing similarities between fish and human embryos were
faked, yet continue to use them as evidence of evolution." [...] Clearly
Haeckel did a dishonest thing with his drawing. Does this mean that the
concept of a phylotypic resemblance among vertebrate classes is a lie? The
answer is a resounding no, and the great indignation raised by Wells is
largely a pious smoke screen. The crucial point is not the superficial
external appearance of embryos, but the sharing of major structural elements
and their topological relationships. (Raff,
2001)
Richardson on creationists and Haeckel.
Richardson himself has been rather annoyed with the creationist response to
his article. Richardson wrote a letter to Science in 1998. His response
applies to Wells equally well:
Our work has been used in a nationally televised debate to attack
evolutionary theory, and to suggest that evolution cannot explain embryology
(2). We strongly disagree with this viewpoint. Data from embryology are
fully consistent with Darwinian evolution. Haeckel's famous drawings are a
Creationist cause célèbre (3). Early versions show young embryos looking
virtually identical in different vertebrate species. On a fundamental level,
Haeckel was correct: All vertebrates develop a similar body plan (consisting
of notochord, body segments, pharyngeal pouches, and so forth). This shared
developmental program reflects shared evolutionary history. It also fits
with overwhelming recent evidence that development in different animals is
controlled by common genetic mechanisms (4).
Unfortunately, Haeckel was overzealous. When we compared his drawings
with real embryos, we found that he showed many details incorrectly. He did
not show significant differences between species, even though his theories
allowed for embryonic variation. For example, we found variations in
embryonic size, external form, and segment number which he did not show (1).
This does not negate Darwinian evolution. On the contrary, the mixture of
similarities and differences among vertebrate embryos reflects evolutionary
change in developmental mechanisms inherited from a common ancestor (5).
[...]
These conclusions are supported in part by comparisons of developmental
timing in different vertebrates (7). This work indicates a strong
correlation between embryonic developmental sequences in humans and other
eutherian mammals, but weak correlation between humans and some "lower"
vertebrates. Haeckel's inaccuracies damage his credibility, but they do not
invalidate the mass of published evidence for Darwinian evolution.
Ironically, had Haeckel drawn the embryos accurately, his first two valid
points in favor of evolution would have been better demonstrated. (Richardson,
1998) Why doesn't Wells quote this letter for his
readers?
The vertebrate classes are not all equally
relatedWells makes much of the embryological differences between the
different vertebrate classes, implying that on evolutionary theory all of the
classes are equally related, but he leaves out the crucial fact that there
is a clear and well-supported consensus about the relationships between the
classes, based on numerous lines of evidence, and that the
embryological development of classes thought to be closely related on
independent grounds are in fact more similar than distantly related
classes, which is the matching independent nested hierarchy pattern that
is predicted by common descent (Theobald,
2002a). Implying, as Wells does, that evolution predicts that all of the
vertebrate classes should be equally similar developmentally is extremely
misleading. Actually, the similarities (taken as characters and put through a
cladistic analysis) should fall into the pattern shown in Richardson et
al.'s second-to-last figure, Figure 9, which of course Wells doesn't show
or mention. Richardson et al.'s Figure 9 is shown here:
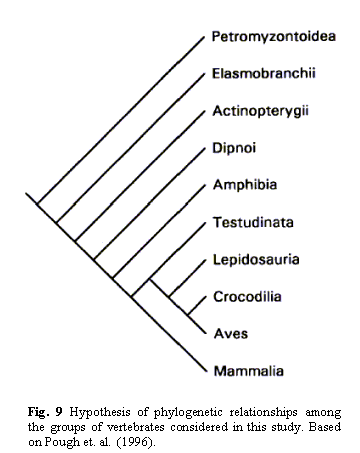
In this figure,
Mammalia are mammals; Aves are birds; Crocodilia, Lipidosauria, and
Testudinata are reptiles; Amphibia are amphibians, and the top four groups are
various kinds of 'fish' (jawed, jawless, etc.). Phylogenetically speaking, all
tetrapods (amphibians, reptiles, mammals and birds) are really just a
sub-subgroup of 'fish.' This figure actually dramatically
underrepresents the massive diversity of 'fish' relative to tetrapods.
For the full view, see the Tree
of Life vertebrates page (for bonus points, see how many steps it takes
you to click through the tree to humans -- if you can find them).
As you can see, mammals, birds, and reptiles are all thought to be more
closely related to each other than any is to amphibians; and mammals, birds,
reptiles, and amphibians together (tetrapods) are all more closely related to
each other than any is to any of the various groups of jawed and jawless fish.
This particular nested hierarchy can be derived from morphology and molecular
phylogeny, but an independent similarity hierarchy based on similarities in
the embryological development pathway also will yield this pattern. See
Theobald for some further discussion: "Ontogeny and Development of Organisms".
- Numerous other mistakes and distortions could be mentioned, here is but
one. Regarding Futuyma's use of the Haeckel embryo drawings in the
3rd edition of Evolutionary Biology, Wells writes (p. 109),
"But it was Futuyma who mindlessly recycled Haeckel's embryos in several
editions of his textbook, until a 'creationist' criticized him for it."
However, an inspection of Futuyma's 1st and 2nd editions
of Evolutionary Biology reveals that no such drawings were
included in these editions. In the first edition, Haeckel's biogenetic law and
the problems with it are discussed on page 153 in respectable fashion (this
corresponds with page 303 in the second edition) -- and in fact the primary
issue surrounding Haeckel in textbooks, which has always been to debunk
Haeckel's "ontogeny recapitulates phylogeny" oversimplification, is in fact
admirably discussed in all three editions.
Archaeopteryx has long been something that creationists have
felt the need to deal with somehow, as it is a clear fossil intermediate between
two vertebrate classes. However, creationist claims have been refuted so often
and so thoroughly regarding Archaeopteryx that very little remains for
Wells to do except raise a smoke screen over whether or not Archaeopteryx
was the actual species through which the genes of the last common ancestor of
modern birds passed, or whether it was a closely related side-branch. Either
way, it is clear evidence that a transition between the classes occurred.
-
It bears pointing out that while Wells makes a big deal about the relative
dating of Archaeopteryx and the various feathered dinosaurs, in fact
the discrepancy is not overwhelming. Archaeopteryx fossils are dated to
be about 150 million years old, and recent work has established a date of 124
mya for the fossils in this area, which puts them in the early Cretaceous (see
the article by Henry Gee at "A date with the downy dinosaurs" ).
Wells claims on pp. 117-122 that cladistics has been used to "rearrange the
fossil evidence" and to provide unwarranted support to the dinosaur-bird
connection. He is greatly troubled by the fact that no feathered dinosaur
fossils earlier than Archaeopteryx have yet been found. Of course,
until very recently we had no feathered dinosaurs at all, which tells you
something about the spottiness of the record for these small,
hard-to-fossilize critters. The total number of known Archaeopteryx
specimens can be counted on two hands, and they are found in a very restricted
locale in Bavaria (although I have recently heard that some new specimens have
been found in Spain). The Chinese sites are similarly restricted as very
special conditions are necessary to fossilize feathers.
-
Wells tries to manufacture another Haeckel embryo situation by describing
the fraudulent fossil Archaeoraptor that National Geographic
foolishly rushed into publication before scientists had a chance to
peer-review the fossil in established journals. But even Wells has to admit
that scientists caught the fraud themselves, that the fossil was never
published in Nature or Science, and that the lifetime of the
fraud was a matter of weeks. A 'Piltdown bird,' to use Wells' deliberately
inflammatory language, this was not. National Geographic was roundly
criticized and will undoubtedly exert more journalistic caution in the future.
Was Archaeoraptor ever in textbooks or even the primary literature? No.
Wells just brings it up to raise the doubt level a bit higher.
-
China has continued to produce magnificent fossils documenting the
dinosaur-bird connection. One wonders what more it could possibly take to
convince one that birds evolved from dinosaurs than the recently discovered
fossil Dromeosaur, which skeletally speaking is entirely a small bipedal
nonflying dinosaur, but which is festooned with feathers of several types?
Excellent pictures of this specimen can be found online at the American Museum
of Natural History website ("New dinosaur specimen" )
and the scientific description is in Ji et al
(2001).
7. Chapter 7: Peppered Moths
So many things are wrong with Wells' treatment of peppered moths (Biston
betularia) that it is hard to list them all; but I will try. The
authoritative reference on this topic is M.E.N. Majerus' 1998 book Industrial
Melanism: Evolution in Action. This book includes two long chapters on
Biston. The first chapter, "The peppered moth story," recounts the basic
story of melanism in Biston, and relates how this story was pieced
together by Kettlewell and others. The second chapter, "The peppered moth story
dissected," gives a thorough critical review of the basic story, considering
aspects and details of the basic story in the light of research (by Majerus and
others) post-dating Kettlewell.
Crucially, however, Majerus clearly and explicitly concludes that, in his
view, Kettlewell got things basically correct. At the beginning of his second
peppered moth chapter, Majerus writes,
First, it is important to emphasize that, in my view, the huge
wealth of additional data obtained since Kettlewell's initial predation papers
(Kettlewell 1955a, 1956) does not undermine the basic qualitative deductions
from that work. Differential bird predation of the typica and
carbonaria forms, in habitats affected by industrial pollution to
different degrees, is the primary influence of the evolution of melanism in
the peppered moth (Majerus, 1998,
p. 116).
Majerus is so clear on this point that one suspects that he was anticipating
that his critique would be misinterpreted by non-peppered moth researchers. It
seems that there is a "too good to be true" quality about the peppered moth
story that leads people to interpret any hint of criticism as a sign that the
whole basic story is crashing down. Scientists are by no means immune to this
tendency, and indeed they may be more prone to it given the regularity with
which popular ideas have been overturned throughout the history of science. The
press has an even greater tendency towards snap judgements and
oversimplifications when it comes to scientific discussions. Antievolutionists,
on the other hand, have always been stuck muttering "it's just microevolution
within a species." While this is true, the rapidity and obvious adaptiveness of
the change effected by natural selection still seemed to give antievolutionists
discomfort. Therefore, it is understandable that when Wells and his fans sniffed
a scientific controversy over peppered moths (in truth it was a fairly marginal
kind of controversy), they blew things way out of proportion.
- First, several of Wells' worst distortions must be
dealt with directly.
-
The natural resting locations of peppered moths -- Majerus' data.
On page 148, Wells discusses the natural resting places of peppered moths,
under the heading "Peppered moths don't rest on tree trunks." But they do,
at least sometimes. Here are the relevant datasets, which Wells does not
quote or cite for his readers:
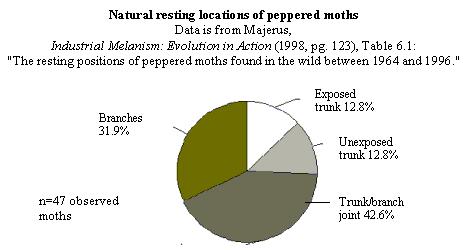
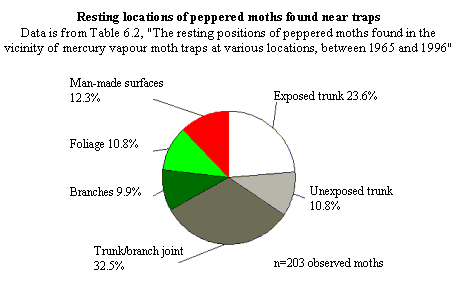
For further discussion, see below and endnote 3.
-
Peppered moth photographs, staged and otherwise. Wells raises a
fantastic stink about the fact that the photographs of peppered moths in
textbooks, showing light-colored typicals next to dark-colored melanics on
differing backgrounds, are staged. But the point of such photos is not
to prove the truth of the 'classic' story, it is to illustrate the relative
crypsis of moth morphs on different backgrounds. Those who feel that
their innocent faith in insect photography has been betrayed should consider
the fact that most photos of insects in textbooks are probably staged; insects
are, after all, small and difficult to photograph. The facts that peppered
moths are sparsely distributed and, well, camouflaged also make them
difficult to photograph.
But as it turns out, the differences between staged and unstaged photos are
minimal. Readers who wish to see unstaged photos of peppered moths are advised
to look up Majerus' Industrial Melanism. Majerus says that all of the
peppered moth photos taken by him in the book are unstaged. Readers should
consult the figures which are listed below. It may be possible to get
permission to include the photos, but until then descriptions shall have to
suffice.
(For those with foggy memories of their texbooks, English peppered moths
come in three general phenotypic categories: typica, the pale, original
'peppered' form of the moth; carbonaria, the almost black melanic form;
and insularia, which includes a range of intermediate-colored
moths.)
-
Figure 6.1 (a), p. 118. Black-and-white photo, edges blurred. A
rather dark (almost black) insularia moth, resting apparently on a
tree trunk (bark fills the background). The moth is slightly darker than the
background.
-
Figure 6.1 (b), p. 118. Black-and-white photo, middle of moth
slightly blurred. A light form of insularia (still more heavily
peppered than a typica), resting on a thick tree branch (branch width
is about 3/4 that of the moth).
-
Figure 6.3, p. 122. Black-and-white photo, middle of moth slightly
blurred. A typica hanging underneath a hazel twig.
-
Plate 3, between pp. 146-147, has colored photos. Six photos are
shown (the first five are Majerus'), and the captions are quoted, with my
comments in brackets.
-
(a) "Typica and carbonaria forms of the peppered
moth on an [sic] horizontal birch branch." [This situation, with two moths
close enough together to photograph at once, is very rare, basically only
occurring if two moths are meeting to mate.]
-
(b) "A pair of peppered moths on a twig at dawn. The
carbonaria male is much less conspicuous than the typica
female." [The carbonaria moth is quite blurry.]
-
(c) "A carbonaria peppered moth in shadow under a
horizontal branch, showing how this positioning may reduce the likelihood
of detection." [The moth is being viewed head-on and is indeed difficult
to see.]
-
(d) "Typical form of the peppered moth at rest during the day in
hazel foliage." [Head-on view, the moth is hanging underneath a thick
twig.]
-
(e) "An intermediate, insularia form, of the peppered
moth." [A 'classic' view, the moth is well-matched to its background,
which is apparently tree bark.]
-
(f) "The non-melanic form of the peppered moth from North
America, Biston betularia cognataria (courtesy of Professor Bruce
Grant)." [A 'classic' view, the moth is well-matched to its background,
which is a lichen-covered surface.]
It should be noted that Majerus is concerned to show his readers aspects of
the peppered moth story that they do not get in textbooks; thus the focus on
insularia forms and on moths in branches (Majerus is a proponent of the
view that peppered moths most commonly -- but not entirely or even
almost entirely -- rest on the underside of branches and thick twigs in the
forest canopy). Even so, there are several photos that show peppered moths, on
tree trunks, on more-or-less matching backgrounds. And guess what? These
photos look no different than 'staged' photos of moths on tree-trunks.
The most 'staged' aspect about a 'staged' photo is that two differing moth
forms are shown side-by-side, but Majerus' first two photos from Plate 3
indicate that even this is not impossible. So the entire photo issue is a
mountain made of a molehill.
It should also be noted that several (four) of these unstaged photos have
some (minor but noticeable) degree of blurring (e.g., part of the moth will be
out of focus). Insects in the wild do annoying things like move and fly away,
and are often encountered in poor-light conditions, resulting in
less-than-perfect photos. As scientific documentation of observations this is
unimportant, but flawed photographs are exactly the kind of thing that are
avoided in textbooks, and this is precisely why staging insect photos is a
common practice for textbooks (as well as things like nature
shows).
Summary of Wells' treatment of moth resting places. To review, Wells'
primary objection to the peppered moth story was this:
Most introductory textbooks now illustrate this classical story of natural
selection with photographs of the two varieties of peppered moth resting on
light- and dark-colored tree trunks. (Figure 7-1) What the textbooks don't
explain, however, is that biologists have known since the 1980's that the
classical story has some serious flaws. The most serious is that peppered
moths in the wild don't even rest on tree trunks. The textbook photographs, it
turns out, have been staged. (Icons, p. 138)
[Figure 7-1 is on Icons, p. 139; these are drawings by
Icons illustrator Jody F. Sjogren; the source photo, if there is one, is
not cited. Confusingly, the caption for the figure is not on page 139 but
overleaf on page 140. These are not encouraging signs in a book purporting to
critique textbooks.]
The discussion thus far has shown that Wells' "most serious objection" to the
peppered moth story is completely baseless: first, peppered moths do in fact
rest on tree trunks (a significant portion of the time although not the majority
of the time, according to Majerus' data). Second, textbook photos are used to
show relative crypsis of moth morphs, not to prove that peppered moths
always rest in one section of the trees. And third, Majerus himself has taken
unstaged photos of peppered moths on matching tree trunk backgrounds, and these
are not significantly different than staged photos; this eviscerates whatever
vestige of a point Wells thinks that he has.
-
What are the implications if moths rest most often underneath
branches? Leaving aside Wells' frantic attempt to create a problem where
none exists, the relevance of moth resting locations for the 'classic story'
(natural selection by bird predation) deserves some consideration. Majerus'
considered opinion is that peppered moths rest more commonly underneath
branches than was previously appreciated, and that if this is true then some
quantitative estimates of selection coefficients may need to be adjusted.
However, he is quite clear that the basic qualitative conclusions of
Kettlewell (that differential bird predation of moth morphs on changing
backgrounds is the selective force) do not need to be changed. As Majerus
notes, crypsis is still important for moths in tree branches. He even comments
directly on this with two of his photos (Plate 3, photos (b) and
(c)). And of course, birds are known to (a) fly and (b) feed in forest
canopies, so it is very difficult to see why resting on trunks vs. branches
would change bird predation in any radical way.
The scientific literature. Having dealt with Wells' "most serious
objection," let us turn to Wells' use of the scientific literature. The primary
problem is that Wells gives inordinate weight to a few scattered review papers,
by biologists who are not major peppered moth researchers [3], that
question the standard view (that bird predation on different colored moths on
differently polluted backgrounds caused the darkening of moth populations as
pollution increased, and that as pollution decreased this process worked in the
opposite direction). Their criticisms have been answered by peppered moth
researchers (Grant,
1999; Cook,
2000; Grant and
Clarke, 2000; Majerus, 2000).
And, as pointed out in the introduction, since Wells bases his argument on the
idea that the experts are disowning the 'icons' in their respective fields,
Wells is falsified if those experts contradict him.
-
Bruce Grant's review of Wells. American peppered moth researcher
Bruce Grant has written many papers on Biston, and has documented the
parallel rise and fall in melanic forms of the North American subspecies of
the peppered moth. See Grant's webpage for
listed articles. Dr. Grant has kindly given permission to have his comments on
the peppered moth chapter of Icons quoted in this article.
To put them in context, the material quoted below is a copy of the
correspondence between Grant and a professional colleague who had requested
Grant's views on Wells' chapter, originally written February 7, 2001.
Subject: Wells' Chapter on Peppered Moths
Wells' Chapter 7 is pretty similar to his earlier ms. "Second thoughts
about peppered moths" that he posted on the web, and published in abridged
form in The Scientist. I sent you my comments about that version
about two weeks ago. My general reaction to this latest version is about the
same. He distorts the picture, but unfortunately he is probably pretty
convincing to people who really don't know the primary literature in this
field. He uses two tactics. One is the selective omission of relevant work.
The other is to scramble together separate points so doubts about one carry
over to the other. Basically, he is dishonest.
He immediately launches the claim "that peppered moths in the wild don't
even rest on tree trunks" (p. 138). This is just plain wrong! Of course they
rest on tree trunks, but it's not their exclusive resting site. He quotes
Cyril Clarke's lack of success in finding the moths in natural settings, but
he omits mentioning Majerus' data which reports just where on trees (exposed
trunks, unexposed trunks, trunk/branch joints, branches) Majerus has found
moths over his 34 years of looking for them. Of the 47 moths he located away
from moth traps, 12 were on trunks (that's >25%). Of the 203 he found in
the vicinities of traps, 70 were on trunks (that's 34%). Based on his
observations, Majerus argued that the most common resting site appears to be
at the trunk/branch juncture. What is clear from his data is that they sit
all over the trees, INCLUDING the trunks. So what? Kettlewell's
complementary experiments in polluted and unpolluted woods compared the
relative success of different colored moths on the same parts of trees in
different areas, not different parts of trees in the same area. It is true
that the photos showing the moths on trunks are posed (just like practically
all wildlife pictures of insects are) but they are not fakes. No one who
reads Kettlewell's paper in which the original photos appeared would get the
impression from the text that these were anything but posed pictures. He was
attempting to compare the differences in conspicuousness of the pale and
dark moths on different backgrounds. Nobody thought he encountered those
moths like that in the wild. At their normal densities, you'd be hard
pressed ever to find two together unless they were copulating. I have always
made a point of stating in photo captions that the moths are posed, and I
think textbook writers have been careless about this. But they are not
frauds.
On the subject of lichens, no one has questioned their importance more
than I have. But what does Wells do with this? He quotes me, but he doesn't
include what else I said has happened on the Wirral (p. 147) with respect to
the tremendous expansion of birch stands since the enactment of the
smokeless zones. Kettlewell, too, argued that peppered moths are well
concealed on birch bark (even without lichens). Wells continues (p. 148) to
quote my reservations about lichens in Michigan, but, again, he omits any
reference to the data I presented in that paper showing the decline, not
only in SO2, but in atmospheric particles (soot) which has been established
as a factor altering reflectance from the surface of tree bark. So, while I
have questioned the importance of lichens, I have not taken this as evidence
that crypsis is unimportant. Wells omits this entirely.
Wells continues to bring up the same old arguments about mysterious other
factors (yet to be identified) that account for the persistence of typicals
in polluted regions, and the presence of melanics in unpolluted locales. He
cites papers written back in the 70s about these puzzles. He omits
discussing in any sophisticated way the role of migration other than to say
"Theoretical models could account for the discrepancies only by invoking
migration...." (p.146), as if in desperation we are forced to grasp at
straws. Of course migration is important. Majerus actually reviews this
point fairly well by comparing the smoothness of clines in melanism between
species that are highly mobile (as is Biston), and species that are
relatively sedentary. Instead of showing his meaningless map of the UK (Fig.
7-2) to illustrate what he regards as anomalies in the distribution of
melanism and lichens, why doesn't he show the before and after comparison
from the national surveys by Kettlewell in 1956, and the survey by Grant
et al. in 1996. (If you'd like, I can send you a jpg file of the maps
I mean.)
Wells also inappropriately uses thermal melanism in ladybirds to suggest,
that while no one has shown this in peppered moths (p. 152), industrial
melanism can have other causes besides predation. It's not just that there
is no evidence for thermal melanism in peppered moths, there is evidence
AGAINST thermal melanism based on the geographic incidence of melanism in
the UK, the USA, and Europe. There are no latitudinal clines, and no
altitudinal clines as one might expect with thermal melanism. Wells knows
this, if he actually read my papers. (He cites them, so I should assume he
read them.) He also raises the question of larval tolerance to pollutants.
There is no evidence for this, either. I have a paper out on this point, but
in fairness to Wells, it came out just this past year.
Wells clouds discussions with irrelevancies. For examples he brings up
Heslop Harrison (p. 141 and again on p. 151) and the question of phenotypic
induction. Wells makes it sound as if most biologists discount induction
based on their belief in natural selection (as if it were a popular
religious question). The evidence for the Mendelian inheritance of melanism
in peppered moths has nothing to do with evolutionary theory; it is based on
old fashioned crosses involving over 12 thousand progeny from 83 broods. The
Mendelian basis for this character in this species is as well established as
is any character in any species. Wells doesn't mention this, yet he cites my
review paper where I do bring this up in my criticism of Sargent et
al. Induction has nothing to do with industrial melanism, and Wells
knows it. Again, selective omissions on the part of Wells.
On page 151 Wells claims Kettlewell's evidence has been impeached. This
is nonsense. It has not. But I have argued, that even if it were entirely
thrown out, the evidence for natural selection comes from the changes in the
percentages of pale and melanic moths. It is this record of change in allele
frequency over time that is unimpeachable. It is a massive record by any
standard. (I can send a jpg file with graphs, if you'd like.) I have pointed
out, and he quotes me, that no force known to science can account for these
changes except for natural selection. Yet, he scrambles the ingredients
here. He claims (top of p. 153) "...it is clear that the compelling evidence
for natural selection that biologists once thought they had in peppered
moths no longer exists." Of course the evidence for natural selection
exists! That evidence is overwhelming. Wells, by attempting to discredit
Ketttlewell's experiments about predation (and clearly there are things
wrong with Kettlewell's experiments) doesn't stop at saying we can't be
altogether sure about bird predation because of problems with Kettlewell's
experiments. No. He says, instead, that the evidence for natural selection
no longer exists. This is just plain wrong. He cannot support this sweeping
statement, but he spins it into his conclusion by building a case against
Kettlewell. This is what I mean about his tactic of scrambling arguments. He
wields non sequiturs relentlessly.
I hope this is helpful to you in your review.
Bruce Grant, Professor of Biology, College of William & Mary.
February 2001
M.E.N. Majerus' review of Wells
-
Majerus, who as we have noted wrote
the (most recent) book on peppered moths and industrial melanism (Majerus, 1998),
wrote a detailed critique of Wells' earlier treatment of peppered moths in
1999. Here is Majerus' opinion of Wells' treatment of peppered moths (in part;
see the links below for the entire correspondence online):
[...]
Evidence of selective predation in the peppered moth is not lacking. It
is just not provided in the quick text book descriptions of the peppered
moth. How can it be. I have read some 500 papers on melanism in the
Lepidoptera. In total, these papers probably amount to about 8000 pages, and
the story is condensed into a few paragraphs in most textbooks for schools.
Even in my own book, I could only give a review of the case covering about
60 pages including illustrations.
The older hypothesis that melanism was induced by pollutants was
discredited because Heslop [Harrison's] experiments lacked appropriate
controls, and his results could not be replicated, despite several attempts.
Furthermore, the levels of mutagenesis that he recorded are several times
higher than those produced by doses of radiation that induce complete
sterility in fruit flies (see E.B. Ford (1964) Ecological Genetics for full
critical review).
Finally, I agree with Dr Wells that photographs of two peppered moths
staged on backgrounds for effect should say they have been done purely for
illustrative purposes. I have many times, in undergraduate lectures, pointed
out that photographs of the type that appear in so many text books are
faked. However, I would point out that none of the photographs of live
peppered moths taken by myself, which appear in the book were staged. All
show peppered moths where they were found in the wild.
End-note: It is difficult to have an informed discussion of a complicated
ecological system with those who have little or no experience of the system.
My advice to anyone who wishes to obtain a fully objective view of this case
is to a) read the primary papers that I based my review upon, and any other
relevant papers, and b) gain some experience of this moth and its habits in
the wild. Of all the people I know, including both amateur and professional
entomologists who have experience of this moth, I know of none who doubts
that differential bird predation is of primary importance in the spread and
decline of melanism in the peppered moth.
I hope that this is some use to you, Donald, and that it encourages more
people to look at the case of the peppered moth with an open mind. If it can
help interest a few more people in moths and butterflies, that is all to the
good.
Best wishes, and Happy Easter.
Mike Majerus
(Majerus, 1999)
-
The above was posted to the Calvin listserv in Donald Frack's detailed
investigation of Wells' claims (e.g. Frank's Peppered Moths -- Round 2).
In his response, Wells freaks out and calls Majerus (who had previously been
Wells' main source for his "peppered myth" claims) a fraud:
BUT EVERYONE, INCLUDING MAJERUS, HAS KNOWN SINCE THE 1980'S THAT PEPPERED
MOTHS DO NOT REST ON TREE TRUNKS IN THE WILD. This means that every time
those staged photographs have been re-published since the 1980's constitutes
a case of deliberate scientific fraud. Michael Majerus is being dishonest,
and textbook-writers are lying to biology students. The behavior of these
people is downright scandalous.
I know what I'm talking about. I spent much of last summer reading the
primary literature (email me if you want the references). Frankly, I was
shocked by what I found -- not only that the evidence for the moths' true
resting-places has been known since the 1980's, but also that people like
Majerus and Miller continue to deceive the public.
Fraud is fraud. It's time to tell it like it is. (caps original, Chadwick's Peppered Moths Again)
Frack says of this,
Note the complete irony of the capitalized sentence. Majerus is the
foremost proponent (in the literature I've seen) of the idea that the moths
most commonly rest higher in the trees. His data are the only ones I have
seen cited as evidence of [what] happens "in the wild." Majerus is attacked
as "dishonest" and "text-book writers are lying to biology students", their
behavior is "scandalous." [...] If Wells is right, he hasn't demonstrated it
here. He attacks both Michael Majerus and Bruce Grant. If Grant's frequent
co-authors, such as Cyril Clarke, are added to the ridicule list (and I
don't know why they wouldn't be), then Wells is well on his way to rejecting
all the well-known researchers on this subject. An awesome, and, at face
value, an incredibly arrogant, claim.
-
There is much more where this came from, and unfortunately there is not yet
a comprehensive web source covering Wells' abuse of peppered moths, so you
have to do some digging. Some good places to start are these links:
-
Bruce Grant's review article: Fine Tunning the Peppered Moth Paradigm
-
Don
Lindsay's archive (links to various letters to newspapers from
Grant and others, protesting Wells' characterizations of their work)
-
NMSR/Wells dialog
-
Wells scored a minor coup when a toned-down version of his essay "Second Thoughts
about Peppered Moths" was published in The Scientist
(13(11), p. 13, May 24, 1999). A longer, unedited version is
here.
-
The 1999 Wells/Frack posts on the Calvin listserv go roughly in this
order (these really should be reformatted and archived somewhere):
-
Frack, "Peppered Moths - in black and white (part 1 of 2)
-
Frack, "Peppered Moths - in black and white (part 2 of 2)
-
Wells, quoted in "Peppered moths again"
-
Frack, "RE: Peppered Moths again"
-
Frack, "Peppered Moths - round 2 (part 1 of 2)"
-
Frack, "Peppered Moths - round 2 (part 2 of 2)"
-
Frack, "Peppered moths, round 3"
-
Frack, "Peppered moths and Creationists"
-
Wells, "My last word"
-
Frack, "RE: My last word"
Of moths and maps
So, the experts disagree with Wells and
furthermore identify just the kinds of deceptive tactics that I have been
talking about. A further instance is Wells' Figure 7-2 (p. 145), a map of
England with four locations marked with letters representing "Discrepancies in
peppered moth distribution." This map deserves enshrinement as exhibit A in
geographer Mark
Monmonier's (1996) book How to Lie with Maps. This is basically
what Wells' map looks like:
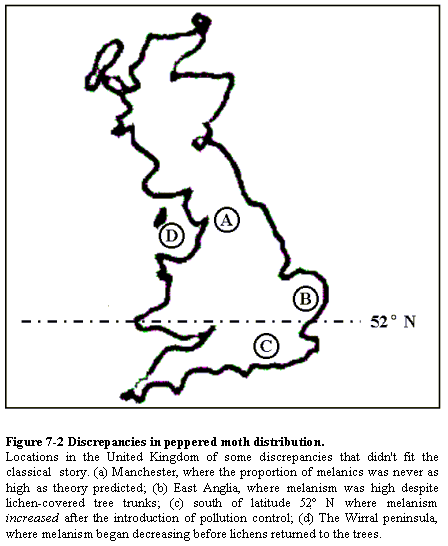
(After Wells,
Icons, p. 145, Figure 7-2. The exact caption is quoted below the
figure. My rendition of the border of Great Britain is very crude, being based
on the first graphic I could find on the internet, but apart from that Wells'
figure is accurately represented. A figure similar to that in Icons can
be found in Wells'
unedited moth essay, here.)
What Wells did here was dig through the literature and find a few instances
where he could find some weak excuse for an observation 'contradicting' what
was expected. However, if one inspects real maps by real moth researchers, one
finds that the geographic pattern is actually a good match with expectations.
Here are the maps that Bruce Grant mentioned in the above-quoted review:
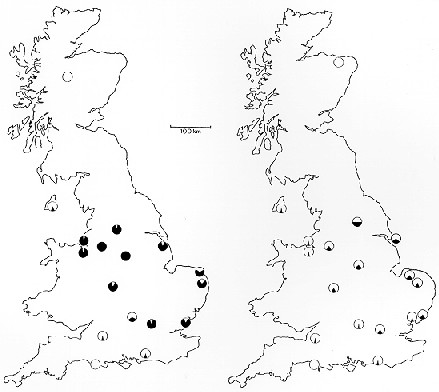
Grant's comments on these maps: "The maps show a before-after comparison of
the geographic distribution of melanic phenotypes in peppered moth populations
in Britain based on Kettlewell's 1956 survey (left map) and that conducted
40-years later (1996) by my colleagues and me (right map). The black segments
of the pie charts indicate the percentage of melanics at the various
locations. Clearly melanism has declined everywhere it was once common."
(Grant, personal communication, February 11, 2002)
The source publication for these maps: Grant, B. S., Cook, A. D. ,
Clarke, C. A., and Owen, D. F. 1998. Geographic and temporal variation in the
incidence of melanism in peppered moth populations in America and Britain.
Journal of Heredity 89:465-471.
Wells' map (Figure 7-2 from Icons) is more fraudulent than all of
the textbook moth photos put together.
8. Chapter 8: Darwin's Finches
"Darwin's Finches" are a group of closely-related bird species located on the
Galápagos islands. The finches have been studied ever since Darwin made the
first collection of these species, but the best-known recent work has been
conducted by the husband-and-wife team of Peter and Rosemary Grant and their
collegues and students. See Peter Grant's 1986 book Ecology and Evolution
of Darwin's Finches and subsequent articles by the Grants, and
especially Jonathan Weiner's 1994 book The Beak of the Finch:
A Story of Evolution in Our Time for a popular introduction to their
work. See also the March-April 2002 issue of American Scientist,
"Adaptive Radiation of Darwin's Finches," by Peter and Rosemary Grant. The
abstract and some graphics are online.
Finches not relevant to Darwin? Wells claims that the finches were not
relevant to Darwin's development of evolutionary theory, but Wells is playing
fast and loose with the facts here. Wells writes (p. 160) that the finches "are
not discussed in [Darwin's] diary of the Beagle voyage except for one
passing reference." Wells (p. 162) does include a quote from the two pages that
Darwin spent on the finches in The Voyage of the Beagle (Wells, like
Weiner, calls it Journal of Researches), but neglects the well-known fact
that this quote constitutes the first publicly published hint of
evolutionary theory from Darwin (Weiner, 1994).
Wells also leaves out mention of the prominent figure that Darwin put in
Voyage of the Beagle, showing the differently-adapted beaks of the
finches. You can read the quote yourself in the online edition of Voyage of
the Beagle.
The various legends that have grown up around Darwin and the finches -- similar
to Galileo and the Leaning Tower of Piza -- are well addressed in Jonathan
Weiner's The Beak of the Finch (p.p. 35-36), minus the patronizing
editorializing of Wells. Darwin made the first scientific collection of the
finches, so the label "Darwin's Finches" is entirely appropriate.
-
The origin of the finches. Wells' figure 8-1, showing the 14 species
of Darwin's finches in a radial pattern, is as usual not very helpful to a
reader interested in the truth. Why does Wells not show the finches in their
traditional classification into four genera (or five or six; see Sato et al.,
2001)?
-
Why, for that matter, does Wells never mention the scientific names of the
finches or their division into multiple genera? Perhaps Wells avoids doing
this because it would undermine his whole argument that the Grants' conclusion
that natural selection (and a few other well-known factors, e.g. isolation) is
responsible for the speciation of the finches is making "more of their work
than the evidence warrants." (p. 174) Wells of course makes no attempt to
propose a better explanation, or even give any reason why their conclusion is
wrong. Wells instead focuses on the back-of-the-envelope calculation of Peter
Grant in a 1991 that under directional selection a speciation event could
occur in 200 years, even though Peter Grant acknowledges in that very
publication that he has observed oscillating selection.
-
Wells pretends that the only evidence for natural selection being
responsible for speciation is the Grants' direct observations. But this is
patently false -- Wells brushes off the extensive character displacement
studies by multiple researchers as "indirect" (p. 164); he notes that
non-oscillating climate or a long-term trend change in the future might make
selection directional, but Wells fails to tell his readers that we have
extensive evidence of just this occurring in the past, e.g. ice ages on the
scale of hundreds of thousands of years; and Wells fails to note that the
oscillating prospects of hybrid finches that the Grants have observed is a
result of competition for resources among individuals of species that
have divided up the available resources by specializing on food sources (this
is character displacement).
-
Wells spends most of the chapter on the Grants' selection and hybridization
studies, but he gives barely a hint of The Main Point of the Darwin's Finches,
namely that overwhelming evidence indicates that these several genera evolved
from a single ancestral species, and that they did so in an adaptive
manner.
-
Wells never stops to think of what might happen if a single finch
population were to arrive on the originally finch-free Galápagos islands.
Currently, the various finch species have evolved to specialize on different
resources (different sizes of seeds, etc.). Thus, the divergence of one
species into two is impeded by prexisting finch species occupying other
niches. But this would not be the case for the first finch population to
arrive at the island. Expansion into new niches would be favored because those
niches would be unoccupied.
-
Parenthetically, Wells completely botches his description of El Niño on p.
168. Wells describes El Niño as "a disturbance in the winter weather patterns
caused by unusually warm air over the Pacific Ocean." ... which is incorrect,
as an El Niño event is actually a change in the sea-surface temperature
gradient across the Pacific. Normally winds push water westwards
along the equatorial Pacific, resulting in the upwelling of cool nutrient-rich
water in the eastern Pacific. When the winds weaken, the warm water "sloshes"
(speaking very colloquially) eastward, resulting in a cooler west Pacific and
a warmer east Pacific, resulting in reduced upwelling (and poorer fishing) in
the eastern Pacific and heavy rains in the Galápagos and in southern
California (see NOAA, 2001 for a
more complete description).
-
An excellent treatment of the finch example is given by Weiner's (1994)
The Beak of the Finch. See also:
9. Chapter 9: Four-Winged Fruit Flies
Wells must have been hard up for icons at this point, considering that in the
text he cites (p. 185) exactly one textbook that uses the example, and
mysteriously leaves this "Icon" out of his textbook evaluation (p. 249). Wells
leaves fossil horses and fossil hominids out of this evaluation, also, although
for different reasons as they are ubiquitously cited in textbooks.
-
Wells' brief (pp. 181-182, see also 245-246) discussion of "biochemical
mutations" (all mutations are biochemical, actually) is another in a long line
of IDist attempts to minimize the importance of things like antibiotic
resistance (and the corresponding importance that such examples confer to
evolutionary biology). Reviews in the mainstream literature, however (e.g. Walsh, 2000) are
never so flippant. Walsh (2000) notes
that bacteria have 'learned' how to pump out antibiotic molecules, destroy
them, and evade them, and discusses several options to new develop new
antibiotics, methods of circumventing bacterial resistance, and strategies for
extending antibiotic lifespan through a more rational use of drugs. A good
discussion of this issue is also found in Weiner's (1994) chapter "The
Resistance Movement."
-
Wells tries (on p. 189) to disqualify all of the above evidence for
beneficial mutations by demanding what he calls "morphological
mutations." Wells brushes off all of the examples of rapid morphological
evolution in the literature (e.g. cichlid mouthpart specialization, Hawaiian
Drosophila, and so forth), as well as the utterly ubiquitous evidence
that changes in genes cause changes in morphology, with a single brazen
hand-wave (p. 190) about how the connection between genetic differences and
morphological differences is "assumed". Wells claims that there is support for
his notions among past biologists and in non-American biologists, but all he
can do in the text is cite a postmodern-sounding argument from a historian. In
the notes he cites a smattering of sources, e.g. Brian Goodwin (no supporter
of extra-genetic inheritance as far as I've heard) and a 1990 article raising
the possibility of somatic inheritance, but Wells doesn't even try to build a
case; presumably he is just trying to up the doubt meter a bit. Wells also
does no serious search for the evidence he says he requires, namely genetic
difference leading to beneficial morphological difference; I, however, was
able to find an example in 30 seconds on PubMed, dealing with the mapping of
genetic loci that correlate with the morphological differences between the
flowers of two closely-related species of monkeyflower (Mimulus) -- one
species is pollinated by bumblebees, the other by hummingbirds, and the 12
specific morphological differences that allow this are each traced to one or a
few loci on the chromosomes (Bradshaw et
al., 1998).
-
The only other evidence that Wells cites in favor of his pet theory about
how not just evolutionary biologists but geneticists and developmental
biologists have got things horribly wrong is an anecdote about a single
lecturer he met at a single meeting in Germany in 1999. Based on the vague
description it sounds like the lecturer was critiquing oversimplified models
of connections between genes and development (and environment, which is also
important), but of course anyone in Evo-Devo is well aware of these kinds of
complexities.
-
I know that the tolerant teacher should operate under the philosophy that
there is no such thing as a dumb question, but Wells manages to ask a
fantastically dumb one on page 192. He asks, "If our developmental genes are
similar to those of other animals, why don't we give birth to fruit flies
instead of human beings?" I don't know if Wells thinks that the Intelligent
Designer is continually intervening to keep organisms reproducing after their
kind or what, but several answers that would occur to anyone who has done any
degree of serious thinking on this issue would include (a) similar genes are
not identical genes; (b) small differences in regulation of genes can result
in big changes; (c) the action of a gene (or the protein from a gene) will
depend on both genetic and environmental context; and particularly (d)
different combinations and arrangements of similar genes can result in very
different patterns. Any serious discussion would consider these blindingly
obvious possibilities in detail.
10. Chapter 10: Fossil Horses and Directed Evolution
It is evident from the chapter title that Wells can find no particular beef
with fossil horses; not only do they not make the list of criteria for textbook
evaluation, they don't even make Wells' list of "10 questions to ask your
biology teacher" (at "Ten questions to ask your biology teacher about evolution by Wells").
There is an obvious question ("Aren't fossil horses pretty darn good evidence
for microevolution adding up to macroevolution"), but it seems unlikely that it
will be added to the list. For a detailed introduction to fossil horses see Hunt (1995).
-
What Wells apparently wants people to believe is that the evolution of
horses was directed. Wells argues that biologists have vigorously tried
to correct the old horse icon (linear evolution) because they are all trying
to prove that evolution is undirected. Of course, all anyone can argue
scientifically about such metaphysical matters is that the pattern doesn't
look like it's going anywhere in particular (just like it doesn't
look like the occurence of a tornado is directed in any obvious way).
Whether evolution is directed, or not directed, in some cosmic sense is not
determinable by science. Wells makes this point when he says that the
hypothesis of directed evolution cannot be disproved by the bush-like pattern
of evidence.
-
Wells, though, tries to have it both ways, keeping his pet notion of
directed evolution unfalsifiable by evidence, but also trying to discern
supporting evidence for directed evolution in the fossil horse record.
-
Wells writes (p. 199), "The mere existence of extinct side-branches doesn't
rule out the possibility that the evolution of modern horses was directed. A
cattle drive has a planned destination, even though some steers might stray
along the way." Icons reviewer Jim Dawson called this passage "an
incredible analogy that evokes an image of God as a cowboy" (Dawson,
2001).
-
Wells, in his figure 10-2 (p. 200) on the modern fossil horse record,
appears to be engaging in the incredibly ironic task of trying to restore an
old, discredited icon of evolution! He leaves out most of the extinct side
branches (and there are very many of those in any real treatment of horse
evolution, see Hunt,
1995), and uses an extra-heavy line to trace the lineage from
Hyracotherium to the modern Equus. In the caption he pens the
following earthshaking observation: "Note that although the new pattern is not
linear, it still shows a continuous lineage connecting Hyracotherium
with the modern horse." Of course, there is a similar lineage connecting
Hyracotherium to every extinct species, also, except that they all went
extinct. In fact, (assuming no special creation of species) every currently
living creature will have a continuous lineage back to its ancestors, and
every extinct creature, well, won't. If Wells writes another book on science,
perhaps he'd like to move into physics and discuss the profound truth that you
can't see the sun at night.
-
On page 202 Wells finally gets around to discussing what is probably the
topic that really motivates him (and the ID movement), namely the supposed
nasty philosophical biases inherent in evolutionary biology. This has been
discussed extensively by others elsewhere and has nothing to do with fossil
horses and precious little to do with textbooks, so for the most part we will
move on. Two points should be made about the words like "random" and
"undirected," which Wells digs out of several textbooks (as if the purpose of
a science textbook is to be a philosophical treatise suitable for word-by-word
analysis for conceivably debatable points).
-
Readers should ask themselves: is the weather "random" and "undirected", or
not?
-
If yes, according to Wells you are subscribing to "materialistic
philosophy in the guise of empirical science" and we should expect Wells to
write his next book about the dogmatic materialism of meteorologists (and
statisticians, etc.).
-
If you think that Wells may be overreacting a wee bit to the implications
of such words (perhaps because of the provocation provided by certain
campaigners for atheism such as Dawkins, who is an excellent zoologist but
who should do a better job of keeping his metaphysics separate from his
science), then join the club, and consider the possibility that evolution is
"random" and "undirected" in the very same way that the weather is
considered "random" and "undirected." (for a reasonable discussion of the
philosophical issues and of the motivations of the ID movement see Pennock,
1999)
Along these lines, it is worth pointing out that the first "random and
undirected" quote that Wells brings up (p. 206) is from a textbook by known
Christian Kenneth Miller, a devout Catholic who is the leading biologist in
the anti-ID movement (see his website and
his book, Miller
(1999), which very sympathetically discusses the religious issues
surrounding the modern evolution debate, including the anti-religious
tendencies of several biologists that IDists love to quote).
11. Chapter 11: From Ape to Human: The Ultimate Icon
At long last, we come to The Ultimate Icon, the one on the cover of
Icons of Evolution: the March From Ape to Man. However, at this
point Wells seems to have completely lost track of textbooks, which you may
recall were supposed to be the topic of the book (this Icon does not appear in
Wells' evaluation criteria on page 249, either). The only mentions of textbooks
in this chapter are listed below:
-
(1) Wells says "Most modern biology textbooks do not even mention Piltdown"
(p. 217).
- (2) Apparently the only thing Wells could find that was juicy enough to
quote in a chapter on 'The Ultimate Icon' was from an interview of Stephen J.
Gould in Raven and Johnson's Biology, where Gould gives his well-known
view that "Humans represent just one tiny, largely fortuitous, and
late-arising twig on the enormously arborescent bush of life." Several points
are worth making:
-
-
Interviews are different from the body of the text, and it is precisely
the place where scientists giving their personal views is perfectly
appropriate.
-
Evaluated as science, the only word which is at all questionable in the
Gould quote is "fortuitous." Interpreted charitably it is even more mild
than "random" and "undirected" that were discussed above. Wells of course
packs all his vitriol towards Gould's philosophy of contingency into that
single word, growling in disgust, "Like so many other things we have
encountered, this is not science, but myth." (p. 228)
-
And finally, Gould's statement is in direct conflict with the ape-to-man
icon that Wells purports to be critiquing in this chapter, and which Wells
put on the cover of Icons of Evolution!
-
It seems likely that Wells couldn't find even a single modern textbook that
contained the "Ape-to-Man" icon -- if they did, Wells would be sure to include
them (it is possible than an occasional textbook includes the icon to
criticize it). I myself distinctly remember my general biology professor
putting up such an "Ape-to-Man" icon, and informing the class in no uncertain
terms that this was an inaccurate and outdated view of human evolution. The
professor then put up some of the numerous spoofs and cartoons of the icon
that she had collected, to make the point, and then put up the modern view,
which is that human evolution follows the branching pattern so common
elsewhere in evolution. Unlike in his horse chapter, Wells makes no attempt to
show readers this new, more accurate, icon.
-
For that matter, Wells spends precious little of this chapter discussing
the actual modern fossil evidence. Pages 212-214 are on Darwin's opinions in
Descent of Man, pages 214-216 are about the interpretation of
Neanderthal fossils in the late 1800's, and pages 217-219 are about Piltdown
man, which it seems no creationist book on evolution is complete without; see
the Piltdown Man webpage for extensive references on the topic.
-
Wells actually gets around to discussing an important fossil skull for a
bit on page 219 (1470, a likely Homo habilis, see "Skull KNM-ER 1470"
for a picture and links), but then hares off for six pages interpreting other
peoples' interpretations of the interpretations of anthropologists of the
hominid fossil record. Nowhere does Wells actually give the reader any
specific idea of what the fossils are like, what their measurements and dates
are, etc. Fortunately, an excellent resource on the web allows you to examine
the pictures yourself and draw your own conclusions (The Fossil Hominids FAQ).
-
Wells (p. 220) quotes Henry Gee writing that all the evidence for human
evolution "between about 10 and 5 million years ago [...] can be fitted into a
small box." But appallingly, Wells somehow fails to inform his readers that
plenty of fossils exist from 5 million years ago onward, which, as Eugenie C. Scott
(2001) pointed out in her review of Wells in Science, "is when
humans evolved." Using Wells' standards, a witness at a murder trial would be
permitted to say that he saw nothing untoward between noon and six, and yet
leave out the fact that he saw a murder at seven.
- Wells focuses on controversies surrounding interpretation, but why worry
about that when you can see the fossils for yourself online and make up your
own mind? Go here: "Prominent Hominid Fossils".
12. Conclusion -- Jonathan Wells' book: Science or Myth?
This concludes the tour of Jonathan Wells' Icons of Evolution.
At the end of Icons, Wells includes an Appendix where he "grades" ten
recent biology textbooks, giving most of them an F (Campbell et al.'s
Biology -- probably the most popular college biology text, by the way --
comes out on top with a D+). Wells then includes "warning labels", which look
suspiciously like cigarette warning labels, which Wells thinks should be put in
textbooks. Has Wells succeeded in making the case which would justify his
harsh judgements? Let us recall what Wells' argument was supposed to be:
"Some biologists are aware of difficulties with a particular icon
because it distorts the evidence in their own field. When they read the
scientific literature in their specialty, they can see that the icon is
misleading or downright false. But they may feel that this is just an isolated
problem, especially when they are assured that Darwin's theory is supported by
overwhelming evidence from other fields. If they believe in the fundamental
correctness of Darwinian evolution, they may set aside their misgivings about
the particular icon they know something about." (Icons, pp.
7-8)
But as we have seen, in every single case, the actual biological experts
in their specific fields of expertise in fact agree that the actual
evidence in their field supports modern evolutionary theory. Furthermore,
many of these scientists have felt sufficiently strongly about this that they
have published critiques of creationist misinterpretations of their work. Many
of these scientists have felt sufficiently victimized by Wells to write specific
rebuttals of him.
Wells might try to argue that he was talking about the "icons" rather than
the general evidence in the field, but still his argument fails. In the cases of
the Miller-Urey experiment, Darwin's tree of life, vertebrate limb homology,
Archaeopteryx, peppered moths, and Darwin's finches, a fair investigation
of the literature has revealed that Wells has no case, and that these "Icons"
are fully deserving of inclusion in biology textbooks. In the cases of the
four-winged fruit fly, fossil horses, and fossil hominids, we discover that
Wells has not even included these cases in his textbook "evaluation" criteria --
perhaps inclusion of these in the criteria would have raised the textbook grades
too much. In any case it is evident that Wells' problems with the four-winged
fruit fly, fossil horses, and fossil hominids are not really with textbooks, but
with extraneous issues -- the real issues in these cases are Wells' bizarre
views about the relationship between genes and development, and his paranoia
that biology textbooks are pushing the view that life is meaningless and
purposeless. I will have a few final words on this subject in a moment, as it is
an oft-recurring theme in antievolutionist writings.
The single "icon" where Wells has some success is with Haeckel's embryos: the
fraction of textbooks that use Haeckel's drawings should replace them with
photographs or more accurate drawings. But even here, the very authority that
Wells cites against Haeckel's embryos, namely M.K. Richardson, has clearly
stated that the actual facts of embryology do indeed support evolutionary
theory, contradicting Wells' interpretation. As Wells' argument is explicitly
based on the views of the experts in their fields, then Wells, to be
self-consistent, would have to concede that he has only scored a point against
certain textbooks, and not against the theory of evolution.
However, let us be generous and grant Wells a full point for the Haeckel's
embryos case. On the other nine "icons," though, Wells has come up empty-handed.
So Wells has earned a 1 out of 10. Even a generous curve would not save Wells
from a flunking grade. One would think that a guy with a Ph.D. from Berkeley
would have done better.
The only thing more discouraging than Wells' grade are the rave reviews that
Wells got from his peers at his current workplace, the Discovery Institute. Wells' fellows at the
Discovery Institute's Center for the Renewal of
Science and Culture lavished praise on Icons of Evolution, with
nary a word of criticism or even mild questioning. Sure, we might expect
antievolutionist demagogues to do their usual braying; for example, in his
typically balanced style Phil Johnson writes of Icons, "This is one of
the most important books ever written about the evolution controversy. It shows
how devotion to the ideology of Darwinism has led to textbooks which are full of
misinformation." But even Michael Behe, who does know some biology and really
should have known better, has apparently become radicalized enough to write,
"Jonathan Wells demonstrates with stunning clarity that the textbook examples
Darwinists themselves chose as the pillars of their theory are all false or
misleading. What does this imply about their scientific standards? Why should
anyone now believe any of their other examples?" This is the same Behe, you will
recall, who accepted the evidence for common descent -- i.e. the tree of life --
as recently as 1996. What is the intellectual status of a movement like
Intelligent Design that cannot bring itself to question even the most outrageous
of Jonathan Wells' distortions? (for the above reviews and others, see
"Reviews of Icons")
I have only had time to refute the major arguments that Wells raises in
Icons; unfortunately this only scratches the surface. A truly thorough
refutation would take a full book, and one rather longer than Icons at
that. I fear that I have not given readers a sufficient impression of just how
deceptive and devious a writer Jonathan Wells is. Through most of the book,
virtually every sentence contains some sort of illegitimate slant,
whether quoting a scientist out of context, or leaving out crucial pieces of
information, or presenting a nonexpert opinion as an authoritative one, or
simply spewing out unsupported exclamations of doubt, derision, and "dogmatic
Darwinism!" Icons is an impressive bit of propaganda, and frankly,
Jonathan Wells is probably the slickest operator that the antievolution movement
has ever produced. His book, packed with quotes and authoritative declarations,
mangling topic after topic in rapid succession, is a calculated attempt to
overwhelm the reader by sheer diversity of material; even the biologically
educated reader is not likely to have the necessary background to spot all of
Wells' tricks. Writing this review required a substantial amount of research and
help from numerous veteran creationism/ID debaters (see Acknowledgements).
But Wells' cleverest move of all was to attack textbooks rather than
taking on the science directly. The all too common response, even from
biologists, has been along the lines of "Well, sure, textbooks have problems,
but this doesn't affect the theory of evolution." This is falling into Wells'
trap. This review has shown that the topics discussed actually do belong in
textbooks, and do constitute good evidence for evolution, according to the
evidence and according to the experts that Wells claims for support. The book
Icons of Evolution is the real scientific travesty. What is Icons really about?
The central irony of Icons of Evolution is that, while
biologists no longer accept and indeed actively debunk the "March From
Ape-to-Man" image on the cover, it appears to be closer to something that
Wells believes. He apparently does not deny common ancestry of humans
with animals; on page 223 (in the middle of six pages of selective quoting about
the subjectivity and disagreements in paleontology) Wells admits, "Obviously,
the human species has a history. Many fossils have been found that appear to be
genuine, and many of them have some features that are ape-like and some that are
human-like." It seems like Wells' next sentence ought to be "Sorry for all the
trouble, folks, I guess I got a little carried away with this book...", but of
course it isn't. As far as anyone can tell, Wells has the idea that "the human
species was planned before life began, and that the history of life is the
record of how this plan was implemented" (see his essay "Evolution and Design").
In other words, to Wells, evolution (with some unspecified touch of ID)
was marching towards a goal of humans, just like the
Apes-to-Man icon on the cover of Icons of Evolution! It seems
likely that the insertion of this metaphysical idea into science education, as
science, is Wells' real goal.
But is it really necessary to force theology into science? As we saw in the
fossil horses chapter, Wells imputes far more metaphysical significance to words
such as "random" and "undirected" than they actually have scientifically.
Scientifically, evolution is described as "random" and "undirected" in the same
way that the weather, earthquakes, and numerous other natural processes are
described as "random" and "undirected." (For that matter, evolution is also
predictable in a way similar to weather and earthquakes.) Does describing the
weather, or evolution, as somewhat "random" really have the offensive
metaphysical implications that Wells thinks?
For an alternative model, we should investigate the central quote of Wells'
last chapter. Wells is severely offended by Dobzhansky's statement "Nothing in
biology makes sense except in the light of evolution" and decries all of the
evil materialist-naturalist metaphysics he sees in it. But Wells, as usual,
fails to give his reader crucial information: Wells fails to say anything about
Dobzhansky's actual metaphysics: Dobzhansky was a life-long Russian
orthodox Christian. Here are some more quotes from the very same article by
Dobzhansky (1973), which is available online at "Nothing in Biology Makes Sense Except in the Light of Evolution".
"It is wrong to hold creation and evolution as mutual exclusive
alternatives. I am a creationist and an evolutionist. Evolution is
God's, or Nature's, method of creation. Creation is not an event that happened
in 4004 B.C.; it is a process that began some 10 billion years ago and is
still under way."
"Does the evolutionary doctrine clash with religious faith? It does not. It
is a blunder to mistake the Holy Scriptures for elementary textbooks of
astronomy, geology, biology, and anthropology. Only if symbols are construed
to mean what they are not intended to mean can there arise imaginary,
insoluble conflicts. As pointed out above, the blunder leads to blasphemy; the
Creator is accused of systematic deceitfulness." (Dobzhansky T., 1973,
"Nothing in biology makes sense except in the light of evolution", American
Biology Teacher 35:125-9)
13. Acknowledgements
This article was composed with valuable help and comments from Bruce Grant,
Bob Hagen, Wesley Elsberry, Michael Hopkins, Burt Humburg, Ian Musgrave, Pete
Dunkelberg, Jesse, igkappa, theyeti, and several others.
[Talk.Origins
Icons of Evolution FAQs]
14. References
Amundson, R. (2001).
"Homology and Homoplasy: A Philosophical Perspective." Produced by Nature
Publishing Group. Accessed online on 1/15/2002.
Baldauf, S. L., Roger, A.
J., Wenk-Siefert, I. and Doolittle, W. F. (2000). "A kingdom-level phylogeny of
eukaryotes based on combined protein data." Science, 290(5493):
972-977.
Bradshaw, H. D., Otto, K.
G., Frewen, B. E., McKay, J. K. and Schemske, D. W. (1998)."Quantitative trait
loci affecting differences in floral morphology between two species of
monkeyflower (Mimulus)." Genetics, V149(N1): 367-382.
Brandes, J. A., Boctor, N.
Z., Cody, G. D., Cooper, B. A., Hazen, R. M. and Yoder, H. S. J. (1998).
"Abiotic nitrogen reduction on the early Earth." Nature,
395(6700): 365-367.
Campbell, N. A., Reece,
J. B. and Mitchell, L. G. (1999).Biology. Menlo Park, Calif., Benjamin
Cummings.
Cavalier-Smith, T.
(2001). "Obcells as proto-organisms: Membrane heredity, lithophosphorylation,
and the origins of the genetic code, the first cells, and photosynthesis."
Journal of Molecular Evolution, V53(N4-5): 555-595.
Cavalier-Smith, T.
(2002)."The neomuran origin of archaebacteria, the negibacterial root of the
universal tree and bacterial megaclassification." Int J Syst Evol
Microbiol, 52: 7-76.
Chyba, C.(1998)."Origins of life: Buried beginnings."
Nature, 395(6700): 329-330.
Cohen, J. (1995). "Novel center seeks to add spark to
origins of life." Science, 270(5244): 1925-1926.
Collins, A. G. and
Valentine, J. W. (2001)."Defining phyla: evolutionary pathways to metazoan body
plans." Evol Dev, 3(6): 432-442.
Conway Morris, S. (1998). The Crucible of Creation:
The Burgess Shale and the Rise of Animals. New York, Oxford University
Press.
Cook, L. M.(2000). "Changing views on melanic moths."
Biological Journal of the Linnean Society, 69: 431-441.
Copley, J. (2001). "The story of O." Nature,
V410: 892-864.
Davis, W. L. and McKay,
C. P. (1996). "Origins of life: a comparison of theories and application to
Mars." Orig Life Evol Biosph, 26(1): 61-73.
Dawson, J. (2001). "Book review: Icons of Evolution --
Science or Myth?" HMS Beagle(97).
Dobzhansky, T. (1973). "Nothing in biology makes sense except in the light of evolution." American
Biology Teacher, 35: 125-129.
Engel, M. H. and Macko,
S. A. (2001). "The stereochemistry of amino acids in the Murchison meteorite."
Precambrian Research, V106(N1-2): 35-45.
Espiritu, D. J., Watkins,
M., Dia-Monje, V., Cartier, G. E., Cruz, L. J. and Olivera, B. M. (2001).
"Venomous cone snails: molecular phylogeny and the generation of toxin
diversity." Toxicon, 39(12): 1899-1916.
Grant, B. S. (1999). "Fine tuning the peppered moth
paradigm." Evolution, 53(3): 980-984.
Grant, B. S. and
Clarke, C. A. (2000). "Industrial Melanism." Encyclopedia of Life
Sciences.
Grant, B. S., Owen, D. F.
and Clarke, C. A. (1996). "Parallel Rise and Fall of Melanic Peppered Moths in
America and Britain." Journal of Heredity, V87(N5):
351-357.
Grant, B. S., Cook, A. D.,
Clarke, C. A. and Owen, D. F. (1998). "Geographic and temporal variation in the
incidence of melanism in peppered moth populations in America and Britain."
Journal of Heredity, V89: 465-471.
Hunt, K. (1995). "Horse Evolution." Produced by
Talkorigins Archive. Accessed online on 1/15/2002.
Ji, Q., Norell, M. A., Gao, K. Q., Ji, S. A. and Ren, D.
(2001).
"The distribution of integumentary structures in a feathered dinosaur."
Nature, V410(N6832): 1084-1088.
Kasting, J. F. and
Brown, L. L. (1998)."The early atmosphere as a source of biogenic compounds."
in The Molecular Origins of Life: Assembling Pieces of the Puzzle. Andre
Brack. Cambridge, Cambridge University Press: 35-56.
Kelley, D. S., Karson, J.
A., Blackman, D. K., Fruh-Green, G. L., Butterfield, D. A., Lilley, M. D.,
Olson, E. J., Schrenk, M. O., Roe, K. K., Lebon, G. T. and Rivizzigno, P.
(2001). "An off-axis hydrothermal vent field near the Mid-Atlantic Ridge at 30
degrees N." Nature, 412(6843): 145-149.
Knoll, A. H. and
Carroll, S. B. (1999). "Early Animal Evolution: Emerging Views from Comparative
Biology and Geology." Science, 284(5423): 2129 - 2137.
Kral, T. A., Brink, K. M.,
Miller, S. L. and McKay, C. P. (1998). "Hydrogen consumption by methanogens on
the early Earth." Orig Life Evol Biosph, 28(3): 311-319.
Majerus, M. E. N. (1998). Melanism: evolution in
action. Oxford ; New York, Oxford University Press, pp. xiii, 338.
Majerus, M. E. N. (1999). "Personal communication to
Donald Frack." Produced by Calvin College. Accessed online on 1/15/2002.
Majerus, M. E. N. (2000). "A bird's eye view of the
peppered moth." Journal of Evolutionary Biology, 13:
155-159.
Miller, K. R. (1999). Finding Darwin's God: a
scientist's search for common ground between God and evolution. New York,
Cliff Street Books, pp. xiii, 338.
Monmonier, M. (1996). How
to Lie with Maps. Chicago, University of Chicago Press, pp. 1-207.
Murakami, T., Utsinomiya,
S., Imazu, Y. and Prasad, N. (2001)."Direct evidence of late Archean to early
Proterozoic anoxic atmosphere from a product of 2.5 Ga old weathering." Earth
Planet. Sci. Lett., 184(2): 523-528.
NOAA (2001). "What is an El Niño?" Produced by NOAA.
Accessed online on 1/15/2002.
Pennock, R. T. (1999). Tower of Babel: the evidence
against the new creationism. Cambridge, Mass., MIT Press, pp. xviii, 429.
Perkins, S. (2001). "New type of hydrothermal vent looms
large." Science News, 160(2): 21.
Peterson, K. J.
and Eernisse, D. J. (2001). "Animal phylogeny and the ancestry of bilaterians:
inferences from morphology and 18S rDNA gene sequences." Evol Dev,
3(3): 170-205.
Raff, R. A. (2001). "The creationist abuse of evo-devo."
Evol Dev, 3(6): 373-374.
Rasmussen, B. and
Buick, R. (1999). "Redox state of the Archean atmosphere: Evidence from detrital
heavy minerals in ca. 3250-2750 Ma sandstones from the Pilbara Craton,
Australia." Geology, 27(2): 115-118.
Richardson, M. K. (1998).
"Letter." Science, 280(5366): 983.
Richardson, M. K.,
Hanken, J., Gooneratne, M. L., Pieau, C., Raynaud, A., Selwood, L. and Wright,
G. M. (1997). "There is no highly conserved embryonic stage in the vertebrates:
implications for current theories of evolution and development." Anat
Embryol, 196: 91-106.
Sato, A., Tichy, H.,
O'HUigin, C., Grant, P. R., Grant, B. R. and Klein, J. (2001). "On the origin of
Darwin's finches." Molecular Biology and Evolution, 18(3):
299-311.
Scott, E. C. (2001). "Icons of evolution - Science or
myth? Why much of what we teach about evolution is wrong, (2000) (English) by J.
Wells." Science, V292(N5525): 2257-2258.
Segre, D., Ben-Eli, D.,
Deamer, D. W. and D., L. (2001). "The lipid world." Orig Life Evol
Biosph, 31(1-2): 119-145.
Theobald, D. (2002a). "29
Evidences for Macroevolution." Produced by TalkOrigins. Accessed online
on 1/20/2002.
Theobald, D. (2002b). "29
Evidences for Macroevolution: A Response to Ashby Camp's "Critique"." Produced by TalkOrigins.
UCMP (2000). "Metazoa: Fossil Record -- Chart of First
Appearances of Metazoans." Produced by University of California Museum of
Paleontology. Accessed online on 10/10/2001.
Von Damm, K. L. (2001). "Lost City found." Nature,
412(6843): 127-128.
Walsh, C. (2000). "Molecular mechanisms that confer
antibacterial drug resistance." Nature, V406(N6797): 775-781.
Weiner, J. (1994). The beak of the finch: a story of
evolution in our time. New York, Knopf: Distributed by Random House, pp. x,
332.
Zimmer, C. (2001). Evolution: The Triumph of an
Idea. New York, Harper Collins.
15. Endnotes
[1] Nic Tamzek is the pen name of Nicholas Matzke.
[2] The introduction to the book, containing a complete list
of Wells' "icons", can be found online
[3] It may be objected here that Sargent (of the paper
Sargent, Millar, and Lambert, 1998, "The "classical" explanation of industrial
melanism: assessing the evidence," in Evolutionary Biology,
30:299-322), is a moth researcher. This is true, but most of Sargent's
work appears to be on moth species other than peppered moths, such as the
genus Catocala. In genera such as Catocala the situation regarding
industrial melanism and selection appears to be different. Majerus barely cites
Sargent at all in his detailed review of peppered moths in Industrial
Melanism: Evolution in Action.
Regarding the Sargent et al. paper, which is the major source for
those proclaiming the downfall of the classical peppered moth story, an
under-realized fact is that it is formally a review of industrial melanism in
moths in general, and in many species other than the peppered moth
(Biston betularia) the evidence is indeed poor that the "classical
explanation" (selective predation against backgrounds changing due to pollution)
applies. Peppered moths of course come up repeatedly as they are the species on
which the most work has been done, and the authors do question the classical
explanation for peppered moths as well. However, the peppered moth experts are
unimpressed, just as they were unimpressed by a 1986 paper by some of the same
authors (Lambert, Millar, and Hughes (1986), "On the classic case of natural
selection", Rivista di Biologia, 79:11-49.). Here is Cook's (2000)
take:
Work by H. B. D. Kettlewell suggested that selective predation was the main
determining factor in B. betularia, and probably in a wide range of other
examples as well. His evidence consisted of surveys which put the correlation
between melanic frequencies and urbanization on a quantitative basis
(Kettlewell, 1958, 1965), demonstration that wild birds would eat the moths if
they found them (Kettlewell, 1955), and the now famous demonstration that
birds discovered most readily the forms least like the daytime backgrounds on
which they rested (Kettlewell, 1973; Rudge, 1999). Colour of resting
background and heterogeneity due to epiphyte cover, appeared to affect
relative visibility. Selective predation became the accepted explanation for
the rise in morph frequency (Majerus, 1998).
Since then, further evidence has been collected. Over the last two decades
industrial environments have become cleaner and melanic frequencies lower
(Clarke et al., 1990; Mani & Majerus, 1993; Grant et al., 1998;
Cook et al., 1999). There has also been some revision of interpretation. It
has been shown that experiments designed to detect and measure selective
predation were carried out in places where moths were not usually likely to
rest if left to their own devices (Mikkola, 1979, 1984; Liebert &
Brakefield, 1987; Grant & Howlett, 1988; Majerus, 1998). There is by no
means a one-to-one relation between reversion of morph frequency and reversion
of epiphyte pattern (Bates et al., 1990; Grant et al., 1998). These findings
cause us to reexamine the story, but they do not obviously require a radical
revision. Estimations of selection appear to show a correspondence between
fitness and frequency. The correlation between the condition of the sites used
in experimental studies and those actually used by the insects is likely to be
high. Further lines of experimentation are suggested, but no previously held
view has been overturned.
The general tone of commentary on Biston studies has, however, altered.
From being treated as a vivid demonstration of natural selection (Luria, Gould
& Singer, 1981, provide an excellent example) and good field
experimentation (Hagen, 1999), the work concerned has come to be viewed with
suspicion (Sermonti & Catastini, 1984; Cherfas, 1987). In a recent review
by Sargent et al. (1998) almost every reference to past work is
predicated by expressions of doubt, reworking ground covered by Lambert et
al. (1986). When discussing predation experiments they conclude ". . .
there seems to be no clear and consistent relationship between the relative
survivorship of different morphs . . . and the frequencies at which the morphs
naturally occur in different environments". Coyne (1998) adopts a similar
tone, saying that the flaws in the work are too numerous to list. This has led
to some alarming reporting, such as Matthews (1999) in the Daily Telegraph
newspaper in Britain, who refers to a "series of scientific blunders" and
states that the experiments are "now thought to be worthless". This article in
turn was linked in its electronic web version to the Creation Science home
page. Recent commentaries are quoted on more than one anti-evolution web site.
A balanced account, which shows the strength of the data in the face of recent
criticism, has been provided by Grant (1999). I propose here to illustrate the
predation results, which Sargent et al. did not do when they criticized
them, and to consider why a radical change in view should have
occurred.
Cook's conclusion about the state of the evidence and the somewhat baffling
criticism the peppered moth story has received:
CONCLUSION
In industrial melanism of Biston betularia, both the original
increase and recent decline in frequency of melanics are striking examples of
natural genetic change closely related to change in the environment. They must
have a selective basis. The experiments demonstrate selective removal. There
is a general correspondence of morph frequency and appearance of backgrounds
likely to be adult resting sites. None of this is in doubt. The evidence is,
however, limited in two ways. First, non-visual components of selection have
not been investigated directly in this species. Analysis of segregating
progenies suggests pre-adult survival differences (Creed et al., 1980) with
carbonaria homozygotes having an advantage over other morphs.
Non-visual selection is certainly indicated in studies of other melanic moths
(Bishop & Cook, 1980), but we have little more idea than Leigh (1911) how
it may operate.
Secondly, the experimental and observational evidence cannot on its own
carry the burden of a particular view of evolution, such as found in Oxford
ecological genetics. Smocovitis (1996) describes how the view of the
[Neodarwinian Modern] Synthesis with which it was associated came to seem
'constricted' to many students of evolution, and to generate a reaction in
favour of more complex models; the last three decades have been a period of
lively debate and controversy. Distrust of the evidence of industrial melanism
may sometimes arise from a wish to question how the example relates to more
complex levels of evolutionary theory. Criticism on these grounds is
misplaced, and can attract the attention of advocates of creationism who see
an evolutionary field in apparent disarray. The Biston story continues
to provide an exceptional opportunity to analyse a pattern of selection. It
should be pursued, along with study of other species with related but
different responses to environmental change.
It should also be pointed out that while Sargent appears skeptical of some of
the evidence for the classical peppered moth story, he has no problem with the
general idea of cryptic (camouflage) coloration in moths; he writes that the
classical explanation is "eminently reasonable", and furthermore he has himself
published on crypsis in moths. A very interesting webpage on Sargent's
observation of the behavior of another moth species, Catocala relicta, is
online.
Notably, the evidence is against this kind of "find a matching
background" behavior in peppered moths. Peppered moths appear to just look for
shaded spots (such as beneath trunk-branch joints), whether or not the moth is
dark or light colored. As Sargent et al. grudgingly acknowledge, this is
actually a good thing for the classic selection-by-bird predation story for
peppered moths, as picking-matching-background behavior would actually
constitute negative feedback that would weaken the effects of natural
selection (i.e., as tree surfaces got darker, the moths would just behaviorally
move to lighter surfaces that matched their camouflage).
Regarding Jerry Coyne's (1998) review of Majerus' book Industrial
Melanism in Nature,
readers should not allow Coyne's excellent reputation as a
nonsense debunker to dissuade them from recognizing that Coyne's review contains
at least one glaring mistake: namely, Coyne writes, "Majerus notes that the most
serious problem is that B. betularia probably does not rest on tree
trunks -- exactly two moths have been seen in such a position in more than 40
years of intensive search. The natural resting spots are, in fact, a mystery."
However, Coyne somehow fails to mention that in the very next paragraph of
Majerus' book (pp. 121-122), Majerus cites his own data on the natural resting
places of moths -- some 47 moths -- not a lot, but far more than two. This data,
listed in Table 6.1 of Majerus' page 123, indicates that peppered moths in fact
rest in diverse locations (12 on tree trunks, 20 on "trunk-branch joints", and
15 in tree branches). Similar percentages are found in Table 6.2 on a larger
dataset of resting moths found near traps.
All of this serves to illustrate that Coyne's opinion on the classical
peppered moth explanation is by no means definitive. Wells, of course, gives
readers very little of the above complexity of the true situation, and instead
quotes the juicy bits of Coyne's review. Wells' own treatment of moth resting
locations appears to be deliberately deceptive. Wells refers to Clarke et
al's (1985) reference to the two moths collected (Clarke was basis for
Coyne's remark also), and goes on to selectively quote two additional papers on
the topic of resting locations, as well as Majerus' book, to support the notion
that moths don't rest on tree trunks. In the notes (p. 304) Wells buries a
reference to Majerus' book amongst the other papers, and refers readers only to
page 116 of Majerus' book, when in fact the section "The resting behaviour of
peppered moths", including the tables, is found on pp. 121-123. Why does Wells
not just put the numbers, which are the best anyone has, in the text for his
audience to read?
This essay first appeared at TalkOrigins.Org

|
|


